CUATRO CIÉNEGAS FISHES: RESEARCH REVIEW AND
A LOCAL TEST OF DIVERSITY VERSUS HABITAT SIZE
W. L. MINCKLEY
Department of Zoology
Arizona State University
Tempe, Arizona 85287
ABSTRACT
The Cuatro Ciénegas basin, Coahuila, Mexico, supports at least 16 native fish species, 8 of which are endemic. Research on this fauna is reviewed. Fishes occupy springs, spring-fed rivers, marshes, playa lakes, ephemeral pools, and artificial canals. Number of species per collection is positively correlated with habitat size in stable springs and spring-fed rivers, but not spectacularly so in unstable natural habitats or canals. Distinctive, different assemblages characterize both stable and fluctuating environments. Ostariophysine and percoid fishes dominate in the former, and cyprinodontoids occupy the latter.
RESUMEN
La cuenca de Cuatro Ciénegas, Coahuila, Mexico, tiene por los menos 16 especies nativas de peces, de las caules 8 son endémicas. Se repasan las investigaciones de dicha fauna. Los peces ocupan manantiales, ríos alimentados por manantiales, pantanos, lagunas playas, charcas efímeras, y canales artificiales. El numero de especies por colecta está correlacionado positivamente con el tamaño del hábitat en manantiales estables y nos alimentados por manantiales, pero no tanto en los hábitats naturales no estables ó canales. Los ambientes estables y los que varían se caracterizan por colectas distintas y diferentes. Los peces ostariofisos y percoides dominan en los ambientes estables, y los cyprinodontoides ocupan los que varían.
INTRODUCTION.-The intermontane basin of Cuatro Ciénegas, Coahuila, Mexico, supports an unusually diverse fish fauna for a North American desert region. Eight families are represented, including both Nearctic (Cyprinidae, Ictaluridae, Centrarchidae, Percidae) and Neotropical (Characidae, Cyprinodontidae, Poeciliidae, Cichlidae) components occupying diverse and abundant aquatic habitats. Fishes of the basin are relatively well known, yet little other than anecdotes has been published on their ecology. The present paper reviews published and unpublished works on the ichthyofauna and quantifies some aspects of species diversity relative to habitat size, providing a local test of patterns often examined on a regional or even worldwide basis.
METHODS AND MATERIALS.-Review of earlier ichthyological studies was from my files of research results from the Cuatro Ciénegas region. Notification that major works were not cited would be appreciated.
Collections of fishes were made by seines, gill nets, and hoop nets, and span the period 1958 through 1981. Clarity and salubrious temperatures of waters in the area make underwater observations both profitable and enjoyable at all times of year, and such were used extensively as an adjunct to direct collecting techniques. Lists of species collected or observed at 46 localities on the basic floor were compiled for a total of 462 samples as follows: 17 headsprings (lagunas and pozos), 191 collections; 10 rivers and creeks (ríos and riachuelos), 127; 11 barrial (playa) lakes, ciénegas(marshes), and ephemeral pools, 83; and 10 canals, 61 samples. Headsprings range from large (>20 ha, > 5 m deep) to small (<10 m2, <10 cm deep), thermal (25 to 380 C) limnocrenes, with linear currents only near inflows and outflows. Rivers and creeks on the basin floor range in size from > 2 m3/sec discharge to a few m3/sec and are spring fed, typically consisting of individual or combined outflows of limnocrenes. Marshes are relatively stable environments most often associated with lagunas, pozos, and rivers. Playa lakes and ephemeral pools are terminal or nearterminal habitats, comprising respectively the sumps for rivers and creeks, and for riparian depressions filled by moderate local rises in water levels. These places experience vast fluctuations in temperature and total dissolved solids (Minckley and Cole 1968), and arc the most severe aquatic habitats present. Canals are mandug channels draining springs or diverting water from rivers. Canalization is the major modification of aquatic habitats in the basin, resulting in lowered water levels in lagunas, with resultant drainage and reduction in size of associated marshlands. Direct modifications were evident in 9 of 18 major headsprings listed by Minckley (1969) in 1958-60: Lagunas Escobeda, Ferrino, Santa Tecla, Tío Candido, Tío Quintero, Anteojo, Orosco, and San Marcos. All of these plus Poza la Becerra were further canalized by 1981; Contreras-Balderas (present symposium) reports further on Poza la Becerra. The area has been occupied by agricultural man since the 16th Century (Alessio-Robles 1938) so some such changes are old; habitats evidencing early modifications were considered stabilized. Thirty-six of the 191 available samples were from modified habitats, and are analyzed separately.
Habitats were ranked from 1 (small) to 10 (large) on a relative basis. Except for playas and ephemeral pools, which varied in size with time and local conditions, and for habitats that have suffered direct modifications, I can detect few changes in discharge of springs and habitats they supply over my 25 years of observations. Some lagunas have enlarged through foundering, new pozos have similarly appeared, and rivers have slightly altered their courses due to lateral slumping or accretion of travertines (see Minckley 1969 for descriptions of processes). The system otherwise appears to have remained stable through time.
Statistical treatments were by least-squares linear regression for correlation of habitat size (rank) and species diversity (species per collection). Species associations were computed as percentage co-occurrence in collections from various habitats.
Research in the Cuatro Ciénegas basin has been under permits to myself and associates from the Mexican Government, and funded largely through National Science Foundation Grants GB2461 and GB647 7-X and personal resources. I thank the many associates who provided assistance in the field and permitted use of unpublished field notes.
RESULTS.-Fishes and Studies of Fishes of the Basin. Endemics dominate the basin ichthyofauna of at least 16 species: Notropis xanthicara (Minckley and Lytle 1969); Cyprinodon atrorus and C. bifasciatus (Miller 1968); Lucania interioris (Hubbs and Miller 1965); Gambusia longispinis (Minckley 1962); Xiphophorus gordoni (Miller and Minckley 1963); Cichlasoma minckleyi (Kornfield and Taylor 1983); and a yet-undescribed species of Etheostoma (Minckley 1969, 1978). Xiphophorus gordoni was relegated to a subspecies of X. couchianus by Rosen and Bailey (1963), followed by Kallman (1964), and discussed further by Rosen and Kallman (1969); Rosen (1979) reversed this Rosen and Kallman (1969); Rosen (1979) reversed this trend, again considering X. gordoni specifically distinct. I also retain it as a full species. I remain convinced that endemic cichlid fishes of the Cuatro Ciénegas basin included in C. minckleyi comprise a species flock (Taylor 1966, Taylor and Minckley 1966, LaBounty 1974, Minckley 1978) rather than a single polytypic species (Kornfield and Taylor 1983), which is to be the subject of another paper. The forms are here treated as a single taxon to reduce further confusion in the literature. Fishes thought to be differentiated at the subspecific level, but yet without formal names, include Dionda episcopa Ictalurus lupus, Lepomis megalotis, and Micropterus salmoides. Astyanax mexicanus, Pylodictis olivaris, Gambusia marshi, Cichlasoma cyanogutattum, and Cichlasoma sp. are undifferentiated or little studied. Cichlasoma sp. in the basin is a variant defined by morphology of its pharyngeal mill (LaBounty 1974) that is otherwise distributed in northern Mexico and southern Texas. The last two forms are combined for convenience as C. cf. cyanogutattum for the remainder of this paper. Gambusia marshi, exists as two distinct color types (Minckley 1962, Arnold 1966), and exhibit variations of 2N = 46 and 48 chromosomes (M. K. Meyer, pers. comm., 1983) in addition to 2N = 42 as reported by Campos and Hubbs (1971). It also inhabits the upper Río Salado system of Coahuila, into which the Cuatro Ciénegas basin now empties, in part as a result of man's activities.
A number of works on Cuatro Ciénegas fishes remain wholly or in part unpublished. Included are behavioral studies of Cyprinodon atrorus by Itzkowitz (1967) (see Minckley and Itzkowitz 1967, Itzkowitz and Minckley 1969), mate-selection investigations of gambusiin fishes by Arnold (1966) that included Gambusia marshi among others, and behavioral ecology of C. atrorus, C. bifasciatus, and their hybrids by Arnold (1972) (in small part Minckley and Arnold 1969). Liu (1969) included Cyprinodon from Cuatro Ciénegas in his comparative studies of behavior in that group (see Liu 1965, Turner and Liu 1977), and Stevenson (1975) analyzed karyotypes of Cyprinodon from the basin and elsewhere (Stevenson 1981). Peden (1970) included G. marshi in an evaluation of courtship behavior and isolating mechanisms in his studies of the genus Gambusia (Peden 1972a-b, 1973). Lysne (1967) studied histocompatibility relative to population size and isolation in Xiphophorus gordoni (see also Kallman 1964). Lytle (1972) detailed taxonomic interrelations of the Notropis proserpinus group of. the middle Rio Grande region, including N. xanthicara from Cuatro Ciénegas, N. rutilus from Rio Salado and adjacent Rio San Juan in northeastern Mexico and others (see also Hubbs and Miller 1978). LaBounty (1974) provided a review of cichlid fishes of northern Mexico, including those from the basin. Smith (1982) treated trophic ecology of Cuatro Ciénegas cichlids.
Other papers dealing with diverse aspects of these fishes include discussions of all or important parts of the fauna by Hubbs and Miller (1965), Taylor and Minckley (1966), Taylor (1966), Minckley (1969, 1978) and Rosen and Kallman (1969). Deacon and Minckley (1974) and Minckley and Brown (1982) included descriptive information on ecology of Cuatro Ciénegas fishes in their treatments of more general subjects. Rivas (1963) and Rosen and Bailey (1963) dealt with taxonomic arrangements of fishes of the genus Gambusia, including both Cuatro Ciénegas species. Minckley (1964) reported on laboratory hybridization experiments involving Gambusia marshi and G. affinis. Cokendolpher (1980) included Cuatro Ciénegas Cyprinodon in his hybridization experiments. Thompson (1979) reported on karyotypes of New World cichilids, including Cichlasoma spp. from the basin. Kornfield and Koehn (1975), Sage and Selander (1975), Kornfield (1981), Kornfield et al. (1982), and Kornfield and Taylor (1983) included data on morphology relative to diet, plus other ecological interrelations, in their studies of Cuatro Ciénegas cichlids. The last authors reviewed mention of these cichlids in other works in their synonymy for C. minckleyi. Jiménez et al. (1981) reported a new genus and species of trematode from the group (see also Guajardo-Martinez, present symposium). Miller (1976, 1978, 1981) compared Cuatro Ciénegas Cyprinodon with their congeners, discussed their evolution and zoogeography, and pointed out problems of survival of the group in general relevant to man's activities in desert areas. Deacon et al. (1979), in dealing with status of certain North American fishes, included a number of Cuatro Ciénegas species as "endangered, threatened, or of special concern," categories applicable to most obligate aquatic organisms in and zones of western North America.
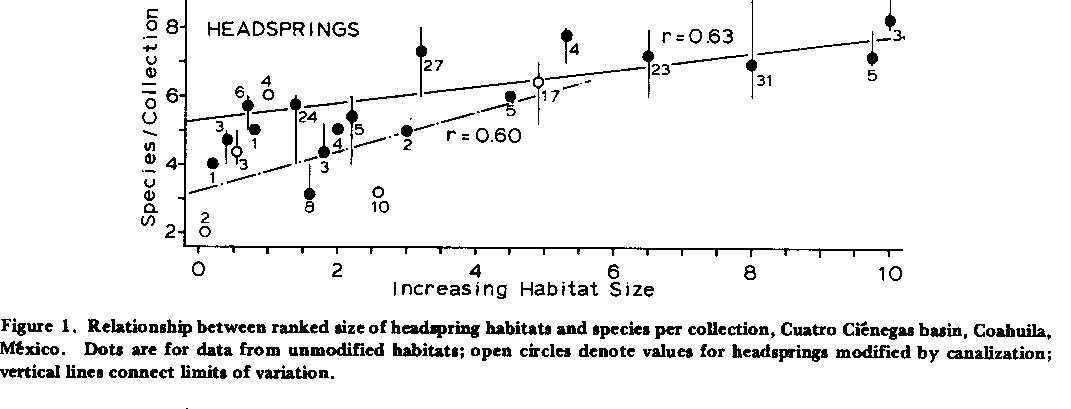
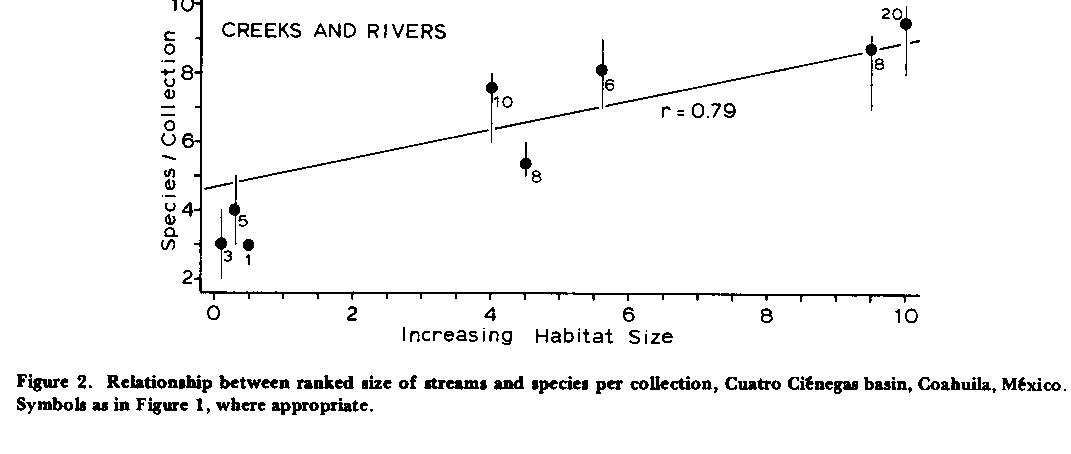
Habitat Size and Fish Diversity. Natural, stable lagunas, pozos, rivers, and creeks of the Cuatro Ciénegas basin yielded more species when large, and fewer species when small (Figs. 1 - 2 ). Correlation between diversity and ranked size is higher for flowing systems than others (r = 0. 79) and appears linear. In unmodified headsprings, smaller habitats have fewer fish species per collection than projected by linear regression, and above an arbitrary rank of ca. 3.0 (>200 M2) seems to add no more taxa (x = 6.9+/-[s.d.] 0.33 species/collection, n = 98; Fig. 1 ) despite a maximum available pool of at least 14 species. Linear correlation between diversity and size-rank for unmodified lagunas (r = 0.63) is not significantly higher (p < 0.05) than that following canalization (r = 0.60), implying that extinction relative to decreasing habitat size is not a factor within observed limits, even when size reduction is artificial.
{kind=link}
{kind=link}
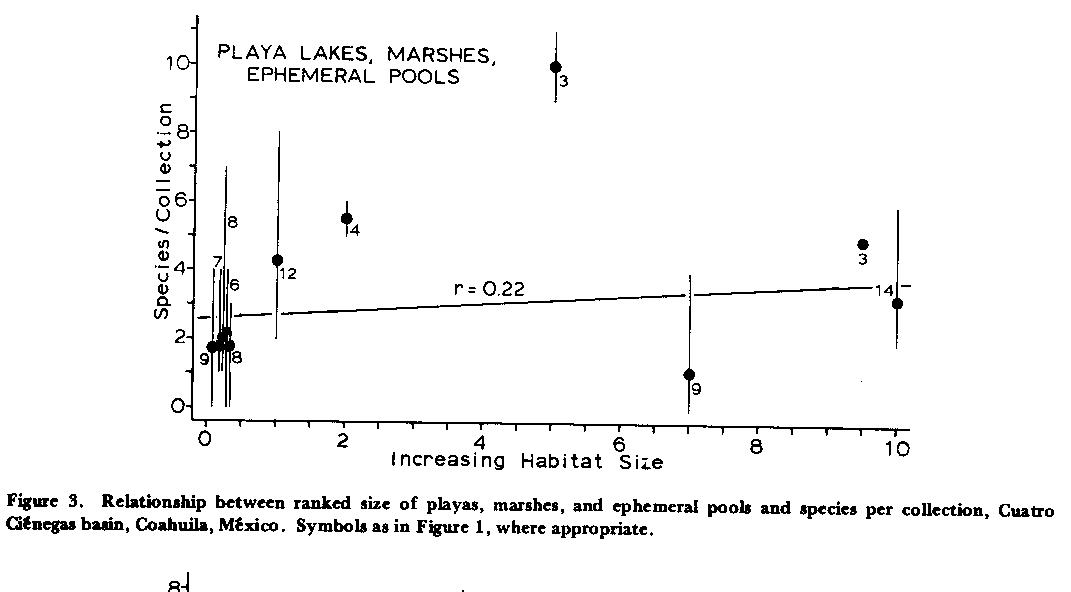
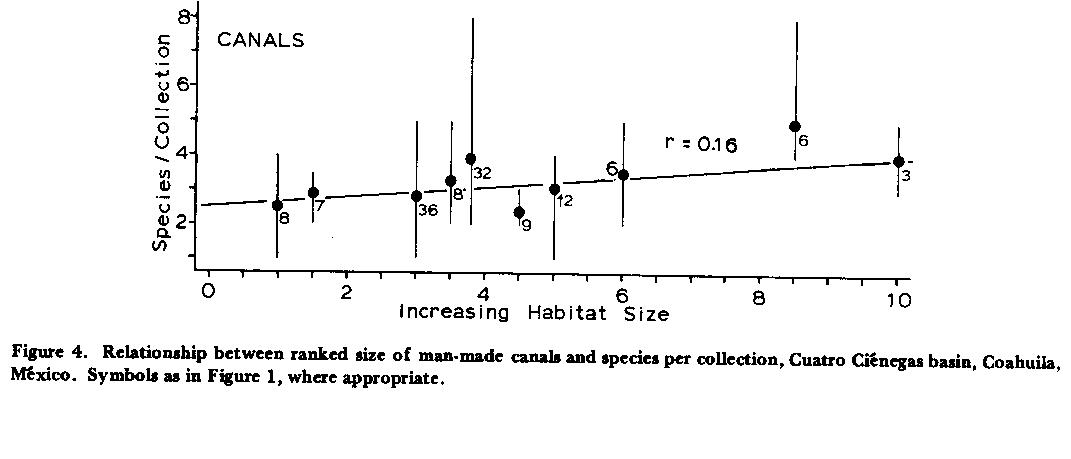
There is little statistical correlation between ranked sizes of playas, marshes, and ephemeral pools and numbers of species (Fig. 3) , but water quality changes select strongly for high salinity and thermal tolerances, thus forcing reductions in diversity as is to be discussed later. Data points for marshes are the highest faunal values in Figure 3, indicating amelioration of severe conditions in those habitats. Canals similarly display essentially no correlation between ranked habitat sizes and fish diversity (Fig. 4) .
{kind=link}
{kind=link}
DISCUSSION.-Measures of habitat size, such as surface area, have long been recognized as correlates of species diversity (Preston 1960, 1962a-b, MacArthur and Wilson 1967). Larger lakes have more fish species than smaller Ones (Barbour and Brown 1974, G. R. Smith 1981). Wider streams, and river or closed basin systems with greater discharges, volumes, or drainage areas Support more kinds of fishes than narrower, smaller volume, lower-discharge systems with smaller watersheds (Hubbs et al. 1974, Contreras-Balderas 1978, Evans and Noble 1979, Welcomme 1979, G. R. Smith 1981, Livingstone et al., Smith and Miller 1984). Similar results in natural, stab of the Cuatro Ciénegas basin thus are not surprising, even with use of size-ranked habitats relative to numbers of species per collection.
The natural laboratory provided by this unique place, its stable habitats at one extreme and markedly unstable habitats at the other, its relatively small size (<1,500 km2 basin surface area), endorheic drainage, lack of relief (basin floor 700 - 760 m elevation), and a relatively large and highly endemic ichthyofauna allow one to avoid or readily interpret some factors that often plague analyses of diversity versus habitat size. Certain features do not exist.
Latitudinal and altitudinal variations that result in gradients of diversity (Fischer 1960, Pianka 1966, Emery 1980) obviously are absent. However, up- to downstream gradients in volume, temporal stability, and spatial heterogeneity, all of which influence fishes (Sheldon 1968, Gorman and Karr 1978, Horwitz 1978, Evans and Noble 1979), are present and well defined. Climatic influences such as seasonal temperature and volume fluctuations are essentially absent in large springs and their outflows, but prevail in downflow habitats. Barriers to dispersal, although present, are minimal on the basin floor. Most systems or system types are or have been interconnected in the recent past (Minckley 1969, 1978; LaBounty 1974), so species richness at a locality could, theoretically, include the entire fauna. A species' absence from a given locale thus constitutes aversion to environmental factors rather than physical exclusion.
Large springs of the Cuatro Ciénegas basin have a distinctive and highly predictable fauna consisting of Astyanax mexicanus, Notropis xanthicara, Ictalurus lupus, Cyprinodon bifasciatus, Gambusia marshi, Micropterus salmoides, and Cichlasoma minckleyi. Spring-fed rivers support the same fishes, plus Etheostoma sp. (Table 1 ). Rheophiles such as N. xanthicara and Dionda episcopa are relatively more abundant in riverline habitats (the latter reaches its greatest relative abundance in canals), and Lepomis megalotis is a more important component of the fauna in rivers. Xiphophorus gordoni also is restricted to headsprings and their immediate outflows; its low frequency of occurrence (Table 1 ) reflects a limited geographic distribution in the system (Miller and Minckley 1963, Minckley 1978). Cichlasoma cf. cyanoguttatum also is restricted in distribution (LaBounty 1974, Minckley 1978).
Species with direct access to, but infrequent or absent in collections from springs and/or rivers, Pylodictis olivaris, Cyprinodon atrorus, Lucania interioris, and Gambusia longispinis, are rare or highly secretive (P. olivaris) or have obvious centers of abundance in other distinctive habitats. Many cyprinodontoids are enhanced by seemingly severe conditions (Miller 1981). Cyprinodon atrorus is remarkably resistant to high temperatures and salinities in desiccating playas and ephemeral pools (Arnold 1972, Deacon and Minckley 1974), and apparently "prefers" such places. It occurred alone in 14 of 64 collections in which it was recorded, and was most commonly associated with the ubiquitous Gambusia marshi (Table 1 ). Lucania interioris and G. longispinis both occupy weedy, ephemeral habitats (Minckley 1962, Hubbs and Miller 1965), and co-occurred there in 64% of recorded collections; G. marshi was typically present with both the last species. Environmental factors in variable habitats of the basin select against or are avoided by other than the cyprinodontoids. Cyprinodon bifasciatus is an exception, being restricted to headsprings, presumably because of its stenothermal tendencies (Miller 1968, Minckley 1978).
Canals were dominated by populations of hybrid Cyprinodon atrorus x C. bifasciatus, Gambusia marshi, and Cichlasoma minckleyi. As noted above, Dionda episcopa was most frequently recorded in canals, and the rheophilic Astyanax mexicanus and Notropis xanthicara were present in these artificial habitats about a third of the time ( Table 1 ).
Data presented here reflect in part collecting techniques and samples available for use from various parts of the basin. Use of gill and hoop nets set overnight at all localities certainly would have increased the number of records for Ictalurus lupus, and possibly Pylodictis olivaris and Lepomis megalotis. Pylodictis olivaris, to my knowledge, has been caught only twice by collectors; other records are visual (underwater) sightings or noted remains of individual fish caught by anglers. Lack of detailed underwater observations definitely reduce numbers of records for species just discussed, and for Etheostoma sp. that lives in interstices of complex, travertine stream-bottoms.
Incomplete field notes or unavailability of notes and/ or specimens for collections made by myself and others also influenced the results. Cichlasoma cf. cyanoguttatum definitely occurs in canals in the eastern portion of the basin (LaBounty 1974, Minckley 1978), but was not included in data sets utilized. Hybrid Cyprinodon similarly occupies spring-fed lakes and streams of the southwestern part of the basin (Arnold 1972, Minckley 1978). Other examples are evident. Future workers dealing with this fauna should examine original literature rather than referring to generalizations such as in this compilation. Graphic presentation of qualitative relations of the fauna by Deacon and Minckley (1974, reproduced by Minckley 1978) are, however, home out by quantitative information on species numbers versus habitat sizes and types and species' associations.
CONCLUSIONS.-Fishes of largely interconnected, springfed aquatic habitats of the Cuatro Ciénegas basin distribute themselves with respect to habitat stability and size. Springheads >200 m2 in surface area and their high-volume outflows have a fauna of ostariophysine and percoid fishes sharply segregated from a cyprinodontoid assemblage in ephemeral pools and playa lakes. Exceptions in springs and spring-fed rivers are Cyprinodon bifasciatus and the ubiquitous Gambusia marshi. Only the latter (abundantly) and Cichlasoma minckleyi of the springhead fauna penetrate terminal waters to any degree, the cichlid only as small individuals in small numbers. Marshes, if deep and associated with springs, provide an intermediate habitat, with, a mixed fauna including components from both ends of the local spectrum. Marshland ichthyofaunas trend toward those of playas when distant from springs and/or shallow and subject to vagaries of direct climatic influence. Springheads, although acting as habitat islands, are inhabited by a predictable fauna of uniform size and composition. In time, long isolation should allow processes of extinction/ colonization to end in clear-cut differences in faunal composition relative to size, heterogeneity, and distances between habitats, but this has not yet occurred.
With natural or man-induced desiccation of such a system, species of transitory and ephemeral (although spring-fed) habitats of the basic floor would presumably be forced toward springs. As springheads decreased in size toward a critical minimum, not specifically defined but perhaps < 200 m2 surface area, extinction processes should prevail. Rheophiles should disappear first, e.g. Astyanax mexicanus, Notropis xanthicara, Dionda episcopal and Etheostoma sp., presumably followed by larger predators (Ictalurus lupus, Pylodictis olivaris, Micropterus salmoides, Lepomis megalotis, Cichlasoma spp.). This would leave cyprinodontoids, a pattern consistent with documented and postulated sequences of extinction resulting in qual- and quantitative faunal changes in other basins of the American deserts, and elsewhere (G. R. Smith 1981, M. L. Smith 1981, Smith and Miller 1985, Minckley et al. 1985).
LITERATURE CITED
ALESSIO-ROBLES, V. 1938. Coahuila y Texas en la Epocha Colonial. Editorial Cultura, Mexico, D. F., Mexico.
ARNOLD, E. T. 1966. Comparative studies of mating behavior in gambusiin fishes (Cyprinodontiformes: Poeciliidae). Unpubl. M. S. 'Thesis, Ariz. St. Univ., Tempe, AZ.
___. 1972. Behavioral ecology of pupfishes (genus Cyprinodon) from the Cuatro Ciénegas basin, Coahuila, Mexico. Unpubl. Ph.D. Dissertat., Ariz. St. Univ., Tempe, AZ.
BARBOUR, C. D. and J. H. BROWN. 1974. Fish species diversity in lakes. Amer. Nat. 108(962):473-489.
CAMPOS, H. H. and C. HUBBS. 1971. Cytomorphology of six species of gambusiine fishes. Copeia 1971(3): 566-569.
COKENDOLPHER, J. C. 1980. Hybridization experiments with the genus Cyprinodon (Teleostei: Cyprinodontidae). Copeia 1980(l):173-176.
CONTRERAS-BALDERAS, S. 1978. Speciation aspects and man-made community composition changes in Chihuahuan Desert fishes. Pp. 405-431, In R. H. Wauer and D. H. Riskind (eds.). Trans. Symp. Biol. Res. Chih. Des. Reg., U.S. Nat. Park Serv. Trans. Proc. Ser. 3 (1977).
DEACON, J. E. and W. L. MINCKLEY. 1974. Chapter VII, Desert Fishes. In G. W. Brown, Jr. (ed.). Desert Biology, Volume II. Acad. Press, Inc., New York, NY.
___, G. KOBETICH, J. D. WILLIAMS, S. CONTRERAS, et al. 1979. Fishes of North America -- endangered, threatened, or of special concern: 1979. Fisheries 4(l):29-44.
EMERY, A. R. 1980. The basis of fish community structure: Marine and freshwater comparisons. Environ. Biol. Fishes 3(l):33-47.
EVANS, J. W. and R. L. NOBLE. 1979. The longitudinal distribution of fishes in an east Texas stream. Amer. Midl.. Nat. 101(2):333-343.
FISCHER, A. G. 1960. Latitudinal variations in organic diversity. Evol. 14(l):64-81.
GORMAN, 0. T. and J. R. KARR. 1978. Habitat structure and stream fish communities. Ecol. 59(3):507515.
HORWITZ, R. J. 1978. Temporal variability patterns and the distributional patterns of stream fishes. Ecol. Monogr. 48(3):307-321.
HUBBS, C. L. and R. R. MILLER. 1965. Studies of cyprinodont fishes. XXII. Variation in Lucania parva, its establishment in western United States, and descriptions of a new species from an interior basin in Coahuila, Mexico. Misc. Publ. Univ. Mich. Mus. Zool 127:1-111, 2pls..
___ and ___1978. Notropis panarcys, n. sp., and N. proserpinus, cyprinid fishes of subgenus Cyprinella, each inhabiting a discrete section of the Rio Grande complex. Copeia 1978(4):582-592.
___, ___ and L. C. HUBBS. 19 74. Hydrographic history and relict fishes of the northcentral Great Basin. Mem. Calif. Acad. Sci. 7:1-259. ITZKOWITZ, M. 1967. Courtship and spawning in a pupfish (genus Cyprinodon) from Coahuila, Mexico. Unpubl. M. S. Thesis, Ariz. St. Univ., Tempe, AZ.
___ and W. L. MINCKLEY. 1969. Qualitative behavior of a pupfish (Cyprinodon atrorus) in differing environments. Grt. Basin Nat. 29 (4)169-180.
JIMÉNEZ, G. F., G. GUAJARDO M., and C. H. BRISENO. 1981. Trematodos de peces dulecacuicolas de Coahuila, Mexico. I. Quadripaludis luistoddi gen. et sp. nov. (Trematoda: Hemiuridae) parasites de ciclidos endemicos de Cuatro Ciénegas. Southwest. Nat. 26(4): 409-413.
KALLMAN, K. 1964. Genetics of tissue transplantation in isolated platyfish populations. Copeia 1964(3): 513-522.
KORNFIELD, I. L. 1981. Biological status of the cichlid fishes of Cuatro Ciénegas. Proc. Des. Fishes Counc. 12:96-97 (abstr.).
___ and R. K. KOEHN. 1975. Genetic variation and speciation in New World cichlids. Evol. 29(3): 427-437.
___ and J. N. TAYLOR. 1983. A new species of polymorphic fish, Cichlasoma minckleyi, from Cuatro Ciénegas, Mexico (Teleostei: Cichlidae). Proc. Biol. Soc. Wash. 96(2):253-269.
___, D. C. SMITH, P. S. GAGNON, and J. N. TAYLOR. 1982. The cichlid fishes of Cuatro Ciénegas, Mexico: direct evidence of conspecificity among distinct trophic morphs. Evol. 36(4):658-664.
LABOUNTY, J. F. 1974. Materials for the revision of cichlids from northern Mexico and southern Texas, U.S.A. (Perciformes: Cichlidae). Unpubl. Ph.D. Dissertat., Ariz. St. Univ., Tempe, AZ.
LIU, R. K. 1965. Evolution of male courtship behavior of fishes of the American genus Cyprinodon, Amer. Zool. 5(4):635-636 (abstr.).
___. 1969. A comparative behavior of allopatric species (Teleostei-Cyprinodontidae: Cyprinodon). Unpubl. Ph.D. Dissertat., Univ. Calif., Los Angeles, Los Angeles, CA.
LIVINGSTONE, D. A., M. ROWLAND and P. E. BAILEY. 1982. On the size of African riverine faunas. Amer. Zool. 11(2):361-369.
LYSNE, J. H. 1967. Histocompatibility studies of the northern platyfish, Xiphophorus gordoni (Poeciliidae). Unpubl. Ph.D. Dissertat., Ariz. St. Univ., Tempe, AZ. LYTLE, G.L. 1972. Revision of the Notropis proserpinus group, subgenus Cyprinella of Notropis, from south Texas and northern Mexico (Pisces: Cyprinidae). Unpubl. M. S. Thesis, Ariz. St. Univ., Tempe, AZ.
MACARTHUR, R. H. and E. 0. WILSON. 1967. The Theory of Island Biography. Princeton Univ. Press, Princeton, NJ.
MILLER, R. R. 1968. Two new fishes of the genus Cyprinodon from the Cuatro Ciénegas basin, Coahuila, Mexico. Occ. Pap. Univ. Mich. Mus. Zool. 659:1-15.
___. 1976. Four new pupfishes of the genus Cyprinodon from Mexico. Bull. S. Calif. Acad. Sci. 75(l):68-75.
___. 1978. Composition and derivation of the native fish fauna of the Chihuahuan Desert region. Pp. 365-381, In R. H. Wauer and D. H. Riskind (eds.). Trans. Symp. Biol. Res. Chih. Desert Reg., U. S. and Mex., U. S. Nat. Park Serv. Trans. Proc. Ser. 3(1977).
___. 1981. Coevolution of deserts and pupfishes (genus Cyprinodon) in the American Southwest. Pp. 39-94, In R. J. Naiman and D. L. Soltz (eds.). Fishes in North American Deserts. John Wiley and Sons, N.Y., NY.
___ and W. L. MINCKLEY. 1963. Xiphophorus gordoni, a new species of platyfish from Coahuila, Mexico. Copeia 1963(3):538-546.
MINCKLEY, W. L. 1962. Two new species of fishes of the genus Gambusia (Poeciliidae) from northeastern Mexico. Copeia 1962(2):391-396.
___. 1964. Hybridization of two species of mosquitofishes (Gambusia, Poeciliidae) in the laboratory. J. Ariz. Acad. Sci. 3 (2):8 7-89.
___. 1969. Environments of the Bolsón of Cuatro Ciénegas, Coahuila, Mexico, with special reference to the aquatic biota. Univ. Tex., El Paso, Cei. Ser. 2:1-65.
___. 1978. Endemic fishes of the Cuatro Ciénegas basin, northern Coahuila, Mexico. Pp. 383-404, In R. H. Wauer and D. Riskind, (eds.). Trans. Symp. Biol. Res. Chih. Des. Reg., U. S. and Mexico. Trans. Proc. U. S. Nat. Park Serv. Ser. 3.
___ and M. ITZKOWITZ. 1967. Ecology and effects of intestinal gas accumulation in a pupfish (genus Cyprinodon). Trans. Amer. Fish. Soc. 96(2): 216-218.
___ and G. A. COLE. 1968. Preliminary limnologic information on waters of the Cuatro Ciénegas basin, Coahuila, Mexico. Southwest. Nat. 13(4):421431.
___ and E. T. ARNOLD. 1969. "Pit digging," a behavioral feeding adaptation in pupfishes (genus Cyprinodon). J. Ariz. Acad. Sci. 5(4):254-257.
___ and G. L. LYTLE. 19 69. Notropis xanthicara, a new cyprinid fish from the Cuatro Ciénegas basin, north-central Mexico. Proc. Biol. Soc. Wash. 82:491502.
___ and D. E. BROWN. 1982. Part 6. Wetlands. Pp. 222-287, 333-341, + Lit. Cited, In D. E. Brown (ed.). Biotic Communities of the American Southwest -United States and Mexico. Desert Plants 4(1-4).
___, D. A. HENDRICKSON, and C. E. BOND. 1985. Geography of western North American freshwater fishes: description and relations to intracontinental tectonism. In press, In C. H. Hocutt and E. 0. Wiley (eds.). Zoogeography of North American Freshwater Fishes. John Wiley and Sons, N. Y., NY. PEDEN, A. E. 1970. Courtship behaviour of Gambusia (Poeciliidae), with emphasis on isolating mechanisms. Unpubl. Ph.D. Dissertat., Univ. Texas, Austin, TX.
___. 1972a. Differences in the external genitalia of female gambusiin fishes. Southwest. Nat. 17(3): 265-272.
___. 1972b. The function of gonopodial parts and behavioral pattern during copulation by Gambusia (Poeciliidae). Canad. J. Zool. 50(7):955-968.
___. 1973. Variation in anal spot expression of gambusiin females and its effect on male courtship. Copeia 1973(2):250-263.
PIANKA, E. R. 1966. Latitudinal gradients in species diversity: a review of concepts. Amer. Nat. 100(l): 33-46.
PRESTON, F. W. 1960. Time and space and the variation of species. Ecol. 41(4):611-627.
___. 1962a. The canonical distribution of commonness and rarity: Part 1. Ecol. 43 (1):185-215.
___. 1962b. The canonical distribution of commonness and rarity: Part II. Ecol. 43(3):410-431.
RIVAS, L. R. 1963. Subgenera and species groups in the peociliid fish genus Gambusia Poey. Copeia 1963(2): 331-347.
ROSEN, D. E. 1979. Fishes from the uplands and intermountane basins of Guatemala: revisionary studies and a comparative geography. Bull. Amer. Mus. Nat. His. 162(5):269-375.
___ and R. M. BAILEY. 1963. The poeciliid fishes (Cyprinodontiformes): their structure, zoogeography, and systematics. Bull. Amer. Mus. Nat. Hist. 126(1):1-176, 2 pls.
___ and K. D. KALLMAN. 1969. A new fish of the genus Xiphophorus from Guatemala, with remarks on the taxonomy of endemic forms. Amer. Mus. Nov. 2379:1-29.
SAGE, R. D. and R. K. SELANDER. 1975. Trophic radiation through polymorphism in cichlid fishes. Proc. Nat. Acad. Sci. 72(11):4669-4673.
SHELDON, A. L. 1968. Species diversity and longitudinal succession in stream fishes. Ecol. 49(l):193-197.
SMITH, D. C. 1982. Trophic ecology of the cichlid morphs of Cuatro Ciénegas, Mexico. Unpubl. M.S. Thesis, Univ. Maine, Orono, ME.
SMITH. G. R. 1981. Effects of habitat size on species richness and adult body sizes of desert fishes. Pp. 125-171, In R. J. Naiman and D. L. Soltz (eds.). Fishes in North American Deserts. John Wiley and Sons, N.Y., NY.
SMITH, M. L. 1981. Late Cenozoic fishes in the warm deserts of North America: a reinterpretation of desert adaptations. Pp. 11-38, In R. J. Naiman and D. L. Soltz (eds.). Fishes in North American Deserts. John Wiley and Sons, N.Y., NY.
___ and R. R. MILLER. 1985. The evolution of the Rio Grande basin as inferred from its fish fauna. In press, In C.H. Hocutt and E.O. Wiley (eds.). Zoogeography of North American Freshwater Fishes. John Wiley and Sons, N.Y., NY.
STEVENSON, M. M. 1975. A comparative chromosome study of the pupfish genus Cyprinodon (Teleostei: Cyprinodontidae). Unpubl. Ph.D. dissertat. Univ. Okla., Norman, OK.
___. 1981. Karyomorphology of several species of Cyprinodon. Copeia 1981(2):494-498.
TAYLOR, D. W. 1966. A remarkable snail fauna from Coahuila, Mexico. Veliger 9:152-228, 11 pls.
___ and W. L. MINCKLEY. 1966. New world for biologists. Pac. Disc. 19(l):18-22.
THOMPSON, K. W. 1979. Cytotaxonomy of 41 species of Neotropical Cichlidae. Copeia 1979(4)-679-691.
TURNER, B. J., and R. K. LIU. 1977. Extensive interspecific genetic compatibility in the New World killifish genus Cyprinodon. Copeia 1977(2):259-269.
WELCOMME, R.I. 1979. Fisheries Ecology of Floodplain Rivers. Longman, London, Engl., and New York, NY.