Environments of the Bolsón of Cuatro Ciénegas, Coahuila, México
With Special Reference to the Aquatic Biota
By
W.L. MINCKLEY
INTRODUCTION
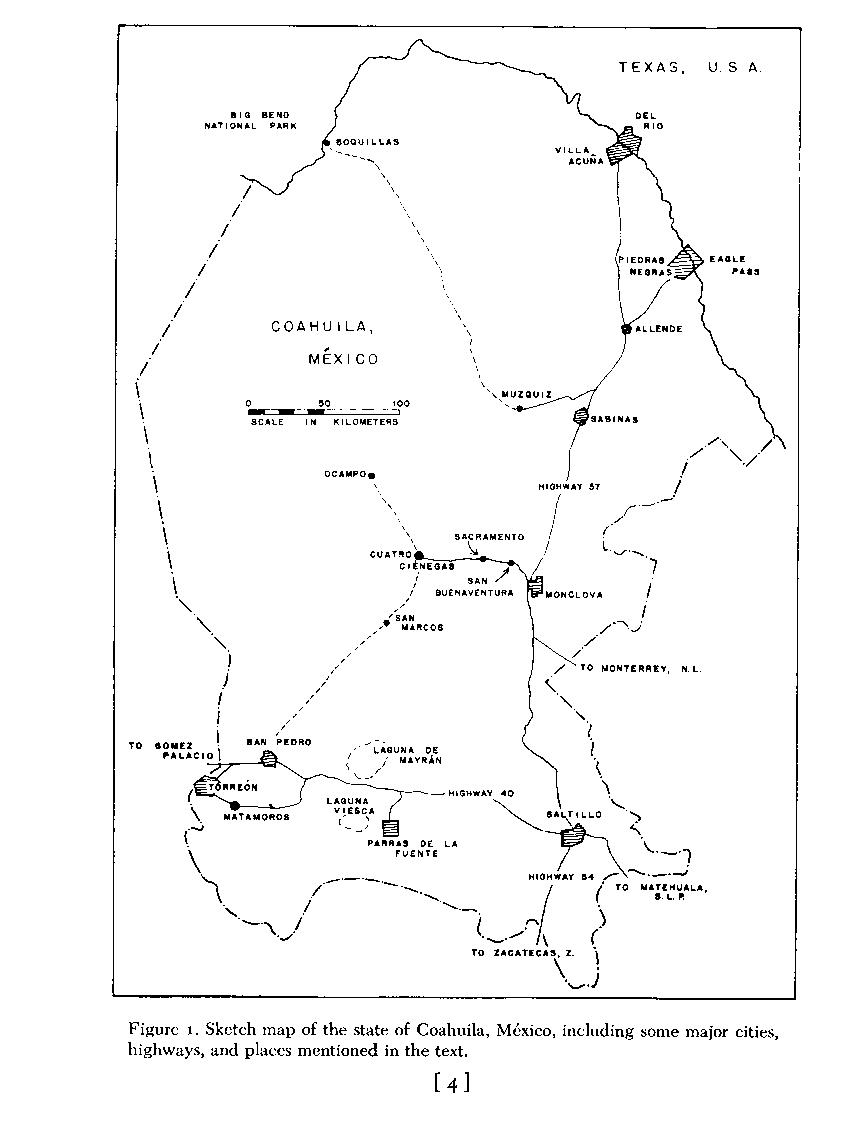
Cuatro Ciénegas de Carranza, hereafter called Cuatro Ciénegas, lies at 26' 59' north and 102' 04' west in the eastern edge of the Mesa del Norte (McBride, 1923; West, 1964), or the Ridge and Basin Province (Humphrey, 1956), of northern México. It is about 270 airline kilometers south-southeast of the big bend of the Río Bravo del Norte (Rio Grande; Fig. 1 ). The small, intermontane valley that lies generally south of the village of Cuatro Ciénegas ( Fig. 2 ) was first explored for biology by E. G. Marsh, Jr., in 1939. His collections form a valuable base for many studies now completed, or in progress. The unique nature of the biota of the Cuatro Ciénegas region was revealed when Carl L. Hubbs examined fishes obtained by Marsh and recognized a remarkably high incidence of endemism (Hubbs and Miller, 1965). Schmidt and Owens (1944) reported on reptiles and amphibians that Marsh caught, including the description of the endemic Coahuilan boxturtle, Terrapene coahuila. It was not until the 1960's, however, that the unique qualities of the fauna were announced in the open literature, and even now a number of remarkable animals remains to be described.
{kind=link}
{kind=link}
In 1958, thanks to John M. Legler, now of the University of Utah, I visited the area. Additional new vertebrates were collected on that trip, and in 1960 intensive studies were commenced. The research proceeded much faster than publication of information, however, especially because of unexpected diversity in the area and the continuing acceleration of disturbance by man's activity. This paper provides data for use elsewhere, principally because of increasing interest in the basin by workers in other disciplines. It results from more than twenty expeditions to the area made by myself, students, and associates in the period from 1958 through 1967. Some information from summer 1968, obtained after this paper was submitted for publication, is included where pertinent. Literature dealing with the area and its biota is reviewed, summaries of data on the various habitats are provided, and interpretations of some aspects of the history and ecology of aquatic and semiaquatic environments are given.
Of special value in a summary of this type, especially to the writer, is the definition of gaps in knowledge -there are many. Some error and misinterpretation seems inevitable; suggestions and additions are therefore eagerly solicited. In those instances where data other than my own are utilized, any errors or omissions are totally my responsibility.
PLACE NAMES
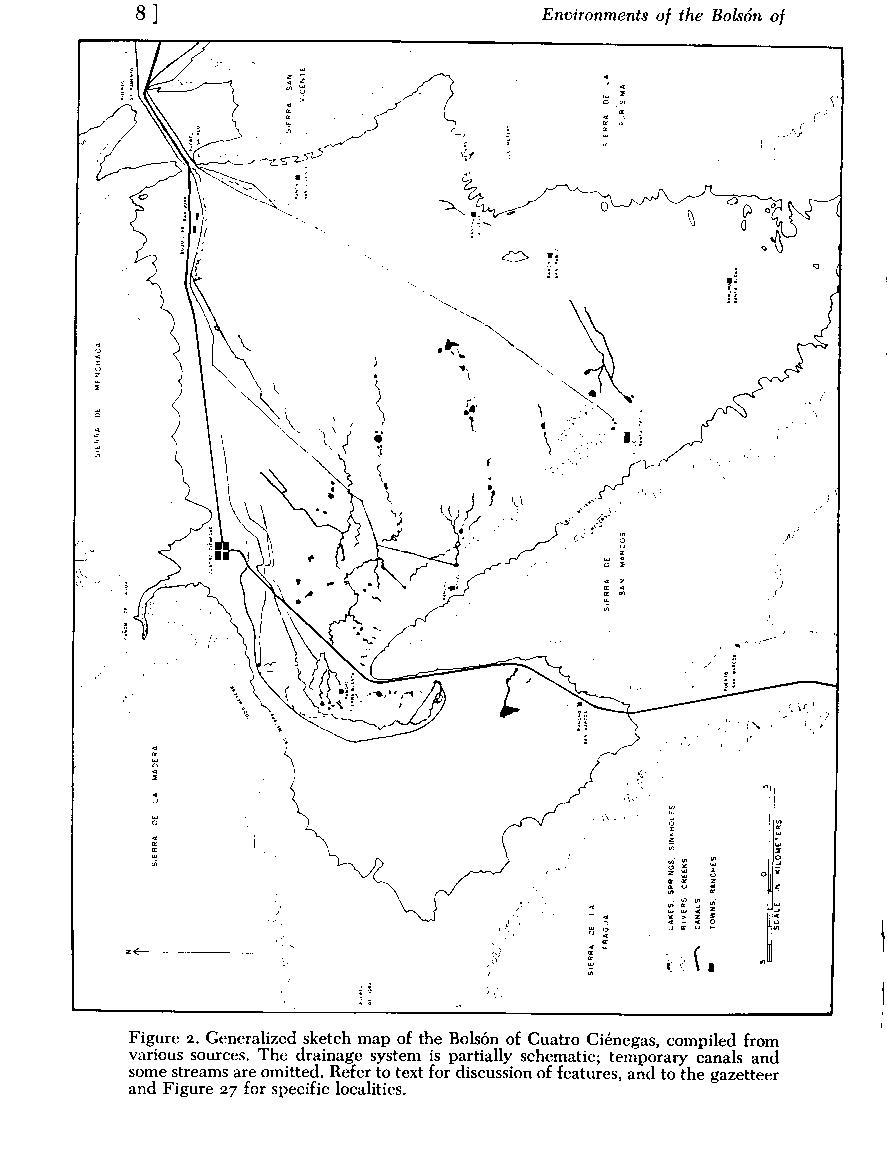
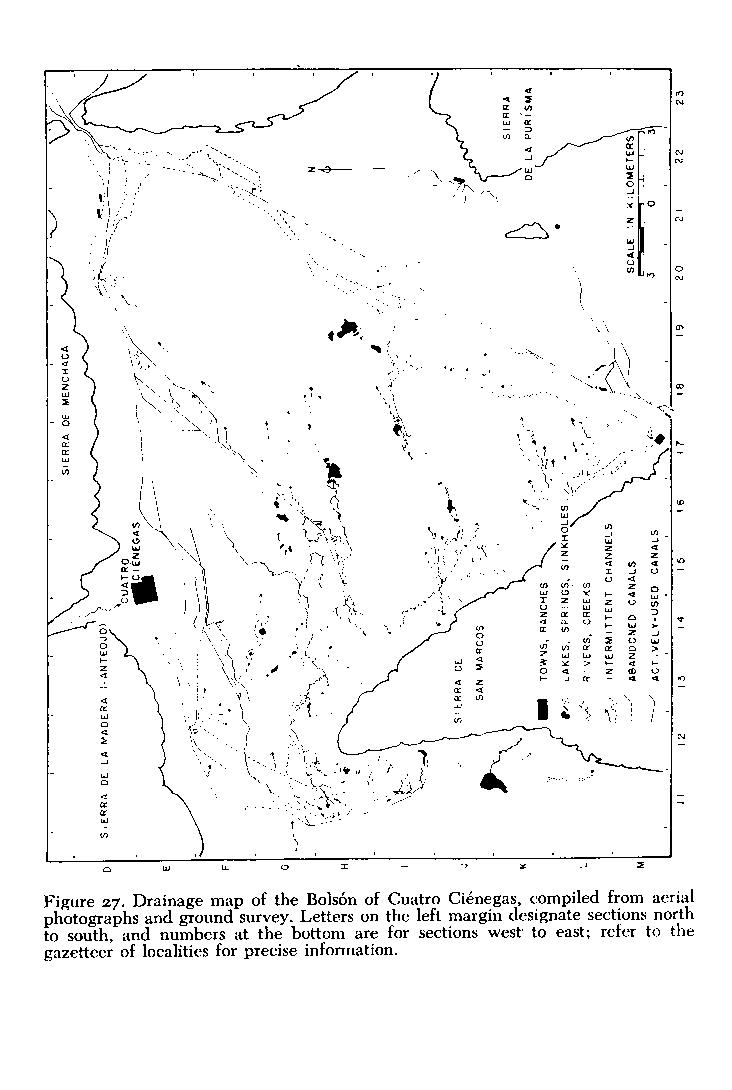
Place names and locality designations are a major difficulty in a compendium such as this, and especially so in the relatively unmapped area of the Mesa del Norte of México. I have provided a sketch map of the basin ( Fig. 2 ), which includes some of the major features. Figures 2 , 3 , 4 and 5 provides a generalized map of the mountains of the area of northern Coahuila where Cuatro Ciénegas is situated. Also, a gazetteer of localities, accompanied by a more detailed map of aquatic systems ( Fig. 27 ) is presented near the end of the paper (P. 50). This last figure is arranged with an approximately 2.5-kilometer grid, letters designating sections north to south, and numbers designating sections west to east. Therefore, the major part of Cuatro Ciénegas is located as "E 15" on that map ( Fig. 27 ). In the text, the various localities are indicated by this code in parentheses following the place name. More precise locations are given by compass subdivision of the square sections (i.e. Ejido Santa Tecla is in W1/2, SE1/4, M 17). This grid is not related to any other system of mapping, except by chance; it was created only for use in this contribution.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Place names in the Cuatro Ciénegas basin may refer to areas rather than to specific lakes or streams; however, some names are specific for a given body of water, and these are marked with an asterisk ( ' ) in the gazetteer. Spelling follows local usage (i.e. the term "pozo," a pit, is locally spelled "poso"); where more than one name or spelling is known, the one most frequently used is favored, and synonyms are in the gazetteer. We have coined some names, and they are given in quotations.
PHYSIOGRAPHY OF THE AREA
Descriptive data on the Bolsón of Cuatro Ciénegas has previously been presented by Gilmore (1947), Legler (1960), Webb and Legler (1960), Minckley (1962), Miller and Minckley (1963), Webb, et al. (1963), Hubbs and Miller (1965), Cole and Minckley (1966, 1968, 1969), Taylor and Minckley ( 1966), D. W. Taylor ( 1966), Boke (1968), Minckley and Cole (1968a-b), and Williams (1968). All but three of these deal primarily with aquatic animals and habitats. An aerial photograph of the basin, from the northeast corner and a high altitude, was published by West (1964), but it is not useful for discussion of specific localities. Additional information on Coahuila may be found in works by Thayer (1966), Gregory (1938), Goldman and Moore (1945),Taylor, et al. ( 1946), Müller ( 1947), Leopold ( 1950), Goldman ( 1951 ), Jaeger (1957), Milstead (1960), Taylor (1962), and papers cited therein.
Physiographic relations of mountains surrounding the basin, and their detrital deposits, are similar to those found throughout the Mesa del Norte (West, 1964) -"The foot of most ranges ... is defined by a sharp angle between the steep mountain face and a more gently sloping rock pediment that grades downslope into the adjacent basin." The floor of the Cuatro Ciénegas basin averages about 740 meters above mean sea level. Sierra de la Madera (locally called Sierra de Anteojo), to the north, rises to more than 3,000 meters and has oaks (Quercus), junipers (Juniperus), and pines (Pinus) on its higher slopes (Baker, 1956). Sierra de San Marcos approaches 2,500 meters at its southern peak, and plunges into the basin floor at its north end (Figs. 2 and 4 ). To the west a low pediment passes through Puerto de Hora. Sierra de la Fragua on the west, and the complex on the east side, range from 1,000 to 1,800 meters in elevation. Mountains on the northeast corner are interrupted by three gaps, in file: Puerto Salado, Puerto Sacramento, and Puerto Nadadores (the last not shown in Fig. 2 ), that now carry an aquatic connection to the Río Grande system.
Dimensions of the basin are about 4o kilometers east to west and 30 kilometers north to south (to an arbitrary limit in the south). Cañon de las Peñas, an extension of the southeast lobe, stretches almost 75 kilometers south-southeast of Cuatro Ciénegas. It has not been explored by zoologists far south of Ejido Santa Tecla (W1/2, SE,1/4, M17).
Just inside Puerto Salado the land surface rises as a broad, alluvial sill interbedded with montane talus debris (Hubbs and Miller, 1965). This sill, which passes across the outlet (northeast) end of the basin, is dissected by two old channels, one from a laguna on the northeast side (D 20, dried between 1958 and 1961), and one from the southeast lobe. Until cut by man-made canals, the sill and other deposits to the west and south, appear to have effectively isolated the central and western parts of the basin, and most of the eastern part, from any external drainage.
Traveling west into the basin the highway passes over alluvium that extends from the Sierra de Menchaca to interdigitate with the basin floor (or barrial). Near Cuatro Ciénegas, an extensive fan from Cañon del Agua (north of D 13-14) extends onto the barrial to meet an area of semi-stabilized dunes about a kilometer south of the town. West of Cuatro Ciénegas the bajada of Sierra de Anteojo is broad, and on the barrial the rocky talus changes abruptly to relatively fine-grained, deep desert soils. The latter represent, in part, an extension of the dune area to about eight kilometers west of town. We traveled 35 kilometers west -the first 15 to 20 were over low desert and bajada along the road and we then climbed gently to the Puerto de Jora over deep soils and talus marked by superficial erosion channels.
Sierra de la Fragua on the west side of the basin exhibits extreme bajada development onto the southwest side of the western arm (see aerial photograph in West, 1964); these appear more extensive than any other alluvial deposit in the area. On the barrial east of this bajada, and extending east almost to the base of the central Sierra de San Marcos, are extensive dunes, positioned variously and in many stages of formation, consolidation, or erosion.
The newly constructed highway leading south from Cuatro Ciénegas crosses the barrial and allows a first impression of the diversity present. It passes through the narrow zone of dunes just south of the village, then through about eight kilometers of relatively flat, slightly depressed desert. The first aquatic habitats are canals in this area. The first natural aquatic habitats are at Puente San Marcos (NW1/4, SE,1/4, G 12), a bridge crossing the Río Mesquites. East and west of the highway are extensive channels and marshes produced by this stream and associated waters. South-southwest of the bridge, ridges of travertine produce local relief to eight meters high; and as one moves toward the tip of Sierra de San Marcos ( Fig. 4 ), large marshes are present on either side of the road.
The highway then traverses the narrow bajada of the west side of Sierra de San Marcos, passing beside the innumerable springs and marshes that lie immediately downslope (Figs 4 and 5 ). About 20 kilometers south, an active dune-field occupies the southwest lobe, as does a lake, Laguna Grande (SE 1/4, J 11, NE1/4, K 11), a few kilometers south and west. From Rancho San Marcos (NE 1/4, SE 1/4, L 11; Figs. 2 , 27 ), the land surface slopes upward toward Puerto San Marcos through which the road leads to San Pedro de Colonias and Torreón ( Fig. 1).
Southeast, along the east side of Sierra de San Marcos, a road crosses numerous alluvial fans and remains generally on rocky baiada for about 20 kilometers. Near Ejido Santa Tecla (W1/2, SE 1/44', M 17), the road reenters the barrial and travels over deep soils that are profusely vegetated with woody shrubs, mostly mesquite (Prosopis). Dense vegetation obscures the terrain; however, it appears to be a gently dissected alluvial fill, similar to, though thicker than that found near the northwest corner and near Puerto Salado. The road along the east side, from near Santa Tecla through Rancho La Vega (W1/2, J 22) and to San Juan (SE1/4, NE1/4, D 21) traverses this kind of sediment through most of its length, west of the bajada of the eastern mountains.
The barrial is complex and certainly not a single unit as the name may imply. This broad, flat area has numerous sinkholes and zones of solution depression, and is dissected in its central parts by the springmarsh-river systems that arc around the tip of Sierra de San Marcos ( Fig. 6 ). This, and other parts of the basin where water occurs, are characterized locally by stony soils marked with ridges and outcrops of marls and travertine-like stone. Such areas are present west, north, and east of the point of the mountain. just south of a line extended from the tip of Sierra de San Marcos to Rancho San Pablo (NW1/4, L 21; Fig. 2 ), development of such relief is pronounced. North of that line the plain is more uniform, with a layer of alkaline soils overlying a calcified basement, and lined occasionally with solidified dune systems.
{kind=link}
A number of playas occur in the central and eastern parts of the basin. Some of these have their central parts filled with water for much of the year, and a few have permanent inflow from spring-fed streams. One playa, Laguna Salada (SW1/4, SW1/4, F 14) just south of Cuatro Ciénegas, is periodically flooded by man for the evaporative collection of salts ( Fig. 6 ), as are three others southwest of Boquillas and San Juan (Southwest of D 21). Playa floors are generally of sparkling white crystalline salts ( Fig. 7 ), and rarely support vegetation of any type.
{kind=link}
CLIMATE
The Cuatro Ciénegas region is climatically and (Müller, 1947). A spread of the "rainy season" from May through October, with a minor average increase in rainfall in December, is indicated by data used by Shreve (1944). The largest number of consecutive rainless months was six, but the periods without rain of three months or more numbered only two in the 20 years of data reported. In no month did the average precipitation exceed 20 millimeters. The area lies within a zone that has a mean annual precipitation of less than 200 millimeters (Contreras Arias, 1942; Shreve, 1944; Vivó Escoto, 1964). As in many desert regions, averages of data on precipitation are deceptive. In 1958 and 1960, rains that exceeded 50 millimeters overnight were encountered, and in 1964 and 1968, more than 30 millimeters of rain fell in four hours. Runoff is rapid from mountains and from some parts of the basin floor. Flashflooding occurs along baiadas, but water usually enters the slopes and little flooding has been observed or is indicated in water-courses of the barrial.
Rains that we have observed are sometimes spectacular. Clouds rise high over surrounding mountains, then move downslope, preceded by strong winds and blowing dust. Winds sometimes persist for days at a time, especially in winter in the southwest lobe of the basin, and act to shape many landforms of the barrial.
Snow occurs on the mountains in winter, according to José Lugo G. (pers. comm.), and occasionally falls within the basin (ca. 75 millimeters in December 1966). Winter temperatures may fall below 0' Centigrade at night, and some desert pools far from spring sources may develop ice that will support the weight of a man. Snow and ice rarely persist in the basin throughout a day.
Air temperatures locally exceed 44.0' Centigrade in summer; however, nights are relatively cool. Variations in air temperature in the shade were from 5.0' to 33.O' Centigrade on a day in April 1963, and a range from 22.5' to 44.0' was recorded in August 1960.
VEGETATION
Vegetation of the area has received little specific study. Johnston (1943, et seq.) reported on collections of plants made in the basin by Marsh, and by himself and colleagues. Gilmore ( 1947) provided specific information on the bajada vegetation. Boke (1968) produced a discussion of some cacti and other plants. In their reviews and summaries, Shreve (1942), Müller (1947), and Cabrera (1955) listed most dominant species and edaphic communities of terrestrial plants in northern México; additional data may be gleaned from general works cited earlier.
The remarkable heterogeneity of desert vegetation, so aptly stressed by Shreve (1942, 1944, and other works), is exemplified in the basin of Cuatro Ciénegas. Rocky soils of bajadas are dominated by low mesquite, creosote bush (Larrea divaricata), tar bush (Flourensia cernua), cbenopods (Suaeda fruticosa and others), and small species of Agave and cacti (many genera). Yucca and various grasses also are common. Mesquite, perhaps because of its conspicuous size, appears dominant throughout the basin wberever deep, well-drained soils persist and there are no travertine or caliche deposits. It is especially abundant on low, stabilized dunes south of Cuatro Ciénegas, and in the southern parts of both lobes. A desert willow (Chilopsis) is locally abundant on less stable dunes. Mesquite, along with infrequent willow (in part Salix nigra), acacia (Acacia greggii), seep-willow (Baccharis glutinosa), and ash (Fraxinus sp.), commonly grow near water. The aquatic and semiaquatic flora is exceedingly rich, and is discussedbelow in the descriptions of aquatic habitats. One group of these, the Charophyta, has been studied (Donald R. Tindall, unpublisbed).2 Transition from bajada to barrial is a change from relatively good drainage and leacbing to poor drainage in fine-grained, highly mineralized, ligbt-colored soils. Here, and locally in more saline areas near playas, Allenrolfea occidentalis (iodine bush), infrequent clumps of bunchgrass (Sporobolus airoides), and salt grasses (Distichlis spp.) are dominant. Consolidated soils of the central barrial are vegetated mostly by Distichlis stricta, but D. spicata and an undetermined species of Distichlis, plus Allenrolfea, also are present. Along the shores of Laguna chenopods occur back from Grande (SE1/4, J 11 , NE1/4, K 11; Fig. 27 ),
the water, while D. stricta lives to the water's edge and is periodically flooded ( Fig. 18 ). Active dunes associated with Laguna Grande ( Fig. 4 ), and elsewhere, have vegetation superficially similar to that described by Shields (1956) for gypsum dunes of the Tularosa basin, New México. Travertine ridges and consolidated dunes in the barrial are vegetated with plants characteristic of bajadas; mesquite, acacia, creosote bush, and an occasional ocotillo (Fouquieria) are conspicuous.
{kind=link}
AQUATIC HABITATS
Surface waters of the Cuatro Ciénegas basin range from ephemeral flooding on playas following rainfall, to large, permanent lakes, rivers, marshes, and springs. Places mentioned in the text are given in the gazetteer of localities at the end of this paper, and may be specifically located on Figure 27 . In some instances, the modifications of the area by man have resulted in changes that have yet to be recognized by change in local names. Thus, a canal along the east side of the basin remains known as Río Polilla, which it partially replaced, and as Canal de Polilla.
Rivers and Creeks - A number of flowing channels occur that are known as "Rios" or "riachuelos." The most extensive is the Río Mesquites, which was originally named the Río San Marcos (Rodriquez Gonzáles, 1926), and has been called variously in recent publications (Webb, et al., 1963, provide clarification). Characterization of this stream serves to describe the basic features of most large watercourses on the barrial.
Immediately southeast of the Rancho Tierra Blanca (N border, NW 1/4, H 12), the major spring of Río Mesquites issues from beneath a stony ]edge as a swift, turbulent rbeocrene. It then receives a number of tributaries from the west and flows between stony banks onto the eastern barrial about due north of the tip of Sierra de San Marcos. In its upper parts, downstream from the swift origin near Tierra Blanca, the stream is 2 to 8 meters wide, ranges to nearly two meters deep, and its current is slow. East-northeast (SW1/4, G 13) of the tip of Sierra de San Marcos the stream passes into an often-subterranean channel that winds through a large marsh; here it is joined by waters from the Mojarral system (SE1/4, G 12, SW1/4, G 13). The river again enters a distinct channel to pass through the barrial. Here it ranges from 2 to 20 meters wide and to 2.5 meters deep (Figs. 8 and 9 ). Width estimates do not take into account bays and inlets along the channel, some of which are more than 40 meters across. Unlike many streams, Río Mesquites has few riffles; deep, swift chutes occur between long, deep, strongly flowing pools. Banks are almost entirely of travertines, in many places undercut for more than two meters, and much of the shoreline is slumped toward and into the water. Large blocks of stone in the channel impede flow and greatly increase diversity of habitat. Bottoms of marl, rubble, and sand prevail in the current, and are of deep, reduced, calcareous muds and plant debris in eddies and bays. A yellow waterlily, Nymphaea, is abundant in quiet areas. Submergent macrophytes such as Chara, Potamogeton, and Utricularia are sparse. Eleocharis and Typha form beds in shallows; the former also grows in channels where current is slow. Steep, undercut banks preclude development of a riparian flora; instead, desert or semi-desert plants live to the water's edge.
{kind=link}
{kind=link}
Water in the Río Mesquites has been crystal clear except following torrential rain when inflow from the immediate banks discolors the water. Currents range from 75 centimeters per second in constricted parts, to essentially none in bays. Discharge is remarkably stable; less than 10 centimeters fluctuation in water levels has been recorded in more than 30 visits to Puente Orosco (Los Corrientes; N border, SE1/4, H 14) due south of Cuatro Ciénegas. Discharges there, estimated by use of Welch's (1948) formula, range from 1.3 to 1.5 cubic meters per second. Calculated discharges in natural channels of the basin almost certainly are too low. This results from flow beneath undercut banks, through lateral surface channels, and movement of water sporadically into and out of underground tubes.
At its downstream end the Río Mesquites now enters man-made canals to flow out of the basin. It formerly drained through intermediate lagunas into a large, closed depression (E-H, 19-21) in the eastern half. According to Rodriguez Gonzáles(1926), the Río de San Marcos (equals Río Mesquites) terminated prior to canalization in the lagunas that gave the town of Cuatro Ciénegas its name. The canal opening the Río Mesquites system to the Río Grande was constructed ca. 1898 (José Lugo, pers. comm.). I
The highest of several water temperatures in the Río Mesquites recorded by me was 31-O' Centigrade at the thermal source near Tierra Blanca, and the lowest was 16.3', in April 1963. Thermal conditions in areas far downstream from the spring sources are undoubtedly more variable than available data indicate.
Observed stabilities of the Río Mesquites are in sharp contrast to conditions in the adjacent Río Puente Chiquito . This smaller stream also has its major origin in a rheocrene (NW1/4, NW1/4, H 15) and flows in a relatively narrow, stony channel, usually passing underground to reappear a short distance to the east as a larger flow. The stream is named for a natural crossing, or puente (S1/2, SE1/4, G 15), south of Cuatro Ciénegas. Fluctuations in levels of water at this puente are extraordinary. On 17 August 1960, the crossing was dry. On the 18th, water flowed across the puente 30 centimeters deep, and on the 23rd, it was 58 centimeters deep and could not be forded by our vehicle. Water flowed 25 centimeters deep on 19 April 1961 ; the next day the puente was dry. In June 1964, August 1967, and June-August 1968, the stream was dry at its downstream end, above the puente, for at least 400 meters - the lowest records to date. Despite rapid fluctuations in water levels, no turbidities have been noted up- or downstream from the crossing.
Banks of Río Puente Chiquito support small riparian trees, shrubs, and forbs locally ( Fig. 10 ). Included are ash, sycamore (Platanus), mesquite, Rhus, and willow. Thick bunchgrasses also occur in areas of fine soils along the banks.
{kind=link}
Land surfaces immediately adjacent to stream channels in the barrial are almost invariably depressed below the level of drier parts of the basin; however, there is no evidence for erosional production of such relief. I interpret this largely as a solution phenomenon within the calcareous substrate. It is probable that reductions in the amount of water beneath the barrial, through natural process or perhaps recently as a result of man, have promoted compaction and sag of the land surface. Dried channels in parts of the basin that have not been directly modified by man, along with lowering water levels in some large lagunas, provide strong evidence for general lowering of the water table.
Rio Cañon (North of D 13-14) is an extremely different stream located in Cañon del Agua north-northwest of Cuatro Ciénegas ( Fig. 11 ). It serves as the domestic water supply for the town, emerging from the ground at il,loo meters above mean sea level and flowing for 5 to 7kilometers to sink into coarse sediments of the canyon floor, enter irrigation channels, or pass into underground storage cisterns. This stream has pronounced pool-riffle development. Riffles are gravelly and range to a meter wide and 10 to 25 centimeters deep in times of normal flow. Pools are 20 to 75 meters long, no more than 2.5 meters wide, and as deep as a meter. Bottoms in pools are silt and sand, with some gravel and boulder near upstream ends. Aquatic plants are common - Chara and Nitella have been found, along with Cladophora glomerata and other algae, Potamogeton, Zannichellia, Nasturtium, and others.
{kind=link}
Rio Cañon is severely flooded on occasion, sometimes with disastrous results in Cuatro Ciénegas. City streets were inundated in 1966, and water entered some homes and shops. At normal discharge the stream is clear, flowing less than 0.25 cubic meter per second. Water temperatures range from 18.0' to 25.0' Centigrade in summer (range observed on a day in August 1960; all subsequent measurements in summer are between those values). In winter, temperatures may be lower than 8.o' Centigrade. The water is richest in carbonates, unlike other waters in the area which are generally richest in sulfates (Minckley and Cole, 1968a). Cañon del Agua supports a rich, varied flora that includes numbers of larger trees. Sycamore, willow, ash, pecan (Carya), and oak are present, plus desert species that occupy more xeric areas along the terraces. Lusb grasses live in areas protected from grazing by livestock, and domestic plants are extensively cultivated near town.
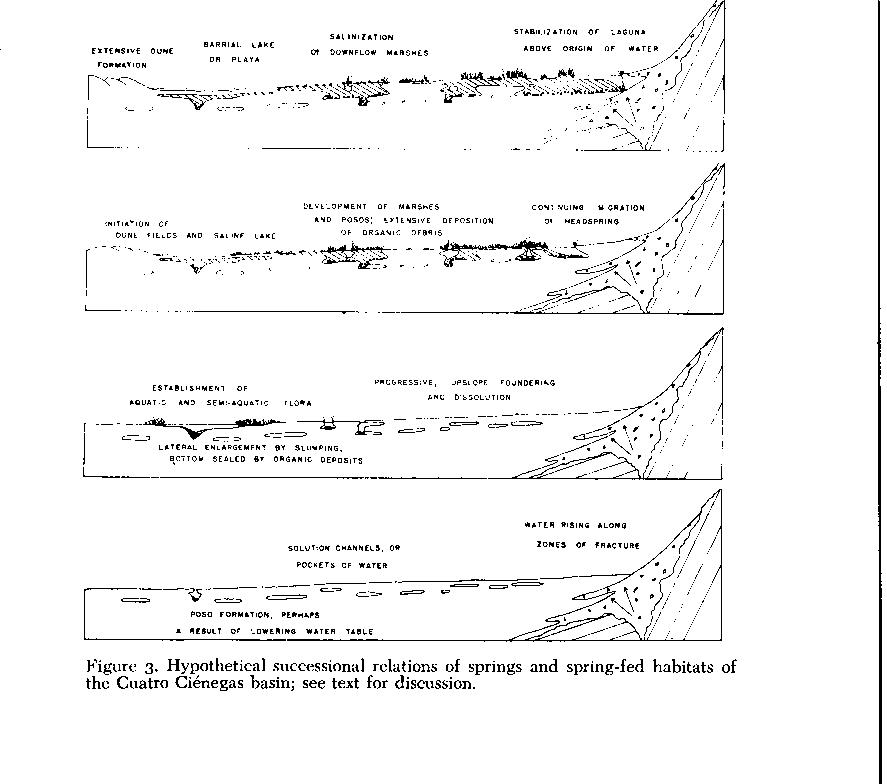
Ponds, Lakes, and Marshes - Natural lentic and semi-lentic habitats in the basin are separable into three major categories. First is a successional series resulting from foundering of subterranean channels, development of resultant pits into lagunas, and a terminal stage(?) of biological succession into marsbes ( Fig. 3 ). A second type of habitat, which may actually consist of an advanced stage of the first category, is represented by permanent, indirectly spring-fed barrial lakes, occupying broad, shallow depressions. And a third type, scarcely studied as yet, is the truly temporary pool that is directly dependent on the vagaries of rainfall.
Habitats of the first series range from a few centimeters in diameter to more than 200 meters in maximum dimensions (excluding associated marsbes), and from less than a meter to more than io meters in depth. Newly foundered pits are locally termed posos, a name that may also be retained for larger lagunas, especially if they have deep, pit-like areas in their bottoms. Numerous recent posos are scattered throughout the basin, some of which have fallen since 1958 and have been observed almost yearly since their origins. Certain parts of the basin have extreme development of karst topography (e.g., in the southeast lobe, Figs. 2 and 27 ). Sinkholes (Figs. 12 and 13 ) often occur in linear series, presumably marking routes of subterranean channels; linearity of depressions in gypsum substrates has been recorded elsewhere (Olive, 1955), and may be a regular feature of such subsidence (Cole, 1963). I have found few dry sinkholes in the basin; current is detectable in the waters of many. Conditions range from active flow from a visible, large tube into one side and out an equal-sized tube on another side - as in the west laguna of El Mojarral (SE1/4, SE1/4, G 12 ), to extremely slow circulation that allows development of some plankton. Plankton has been noted in only three of more than a hundred posos that I have visited. Some subterranean outflows are large enough to necessitate cautious approach by a swimmer; persons unfamiliar with the area should exercise care in their exploration.
{kind=link}
{kind=link}
Conditions in posos of Cuatro Ciénegas can perhaps be compared with those described by Pearse (1936) for cenotes of the Yucatdn Peninsula. Slow currents, at best, were found there. Most appeared cut off from ground-water percolation, and showed chemical and thermal stratification; such is not the case in Cuatro Ciénegas. Cole's (1963) description of Montezuma Well, Arizona, and Navarre's (unpublished) data on Bottomless Lakes, New México, correspond in many ways with situations seen in larger posos and lagunas of the Cuatro Ciénegas region.
A newly foundered poso may undergo a number of alternative successional changes ( Fig. 3 ). A pit, after initial enlargement by slumping of its walls until angular stability is achieved, may remain relatively unchanged for a long time. If active, percolative communication is maintained with the water table, as opposed to strong flow through the pit or to plugging of either the in- or outflow, a rim of aquatic vegetation may form. Where water levels and movements remain constant, semi-floating sedge mats develop. These may actually close over the water surface, and with geochemical influences, described later, may re-roof the poso and convert its surface to dry land. If the inflow is plugged or water levels recede, stagnation and successional filling occurs. Some circular, depressed, almost-dry marshes in the basin document this terminal stage. If an outflow is plugged, and inflow persists, water may rise over the land surface in an epigean flow. Few erosion channels are formed, however, perhaps because of development of marsh vegetation, or because of deposition of salts or travertines that exceeds erosional competence of the water. Desert insolation and evaporative rates enter the picture, sometimes resulting in formation of "cone springs" like those found in New México (Harrington, 1948), and elsewhere. Escobeda (E1/2 , NW1/4, I 14; Fig. 14 ), a large, circular, warm spring, overflowed its banks before canalization to form pronounced travertine ridges.
{kind=link}
Another alternative, differing greatly from the formation of small, stabilized posos, produces the deep, tortuous stream channels in the basin, and ultimately forms the striking spring-fed lagunas (Figs. 15 and 16 ). Development of these complex lake-springs begins with sinkhole formation. Subsequently, in an actively-flowing aquifer-system, foundering and perhaps dissolution of the banks produce a linear, tortuous channel. In systems on the barrial, far from the mountainfronts, continuing headwater foundering probably produces elongate channels similar to that occupied by the Río Mesquites.
{kind=link}
{kind=link}
Lateral enlargement of such systems may occur by downstream overflow, or by collapse of the banks, with increases in area accompanied by increasing heterogeneity and decreasing speed of flow. Accumulation of allo- and autochtbonous debris lateral to and in downstream ends of channels is balanced by the physical competence of water for removal, and with reduction in currents, water-carried debris tends to fill, and block, subterranean outflows. In deserts, terminal stages in succession at downflow ends of a system of this type may be dominated by chemical factors. Waters in desert regions are often highly mineralized, as they are in the Cuatro Ciénegas area (Minckley and Cole, 1968a; Cole and Minckley, 1968), and salts are deposited quickly and in abundance. Terminal parts of most large systems in the basin consist of shallow barrial lakes, or playas, in most cases (Figs 4 , 6 - 7 , 17 - 18 ). Fertile swales of grass, sedge, or other plants persist only where substantial flows of water are present.
{kind=link}
Lagunas occur mostly along the mountain fronts, with the upstream ends nearest the mountains at the downslope terminations of bajadas. I assume that waters of the Cuatro Ciénegas basin rise largely along fracture zones associated with structural features of the mountains ( Fig. 3 ). Therefore, upflow development of laguna channels ends at the point of vertical origin of water. The mountain mass, and filling of headwaters by montane debris, no doubt also stop upstream erosion. With such a finite point of headward progression, assuming relatively constant inflow of water, steady-state conditions may have prevailed in the upper parts of the lagunas for millenia.
Posos, streams, and lagunas are greatly modified by salt deposition. Development of sedge mats at the water's edge provide, through death and accumulation, substrate for development of additional vegetation - a classical sequence. However, encroachment by plants on the water side is accompanied by salt accumulations on the landward side as a result of evaporation of water drawn up by capillarity from the spongy sedge-peat. Salts are deposited at the surface in ever-increasing thickness, while the least soluble components (principally calcium carbonate) form a travertine-like level beneath more soluble, and more toxic, salts, such as sodium chloride. The last accumulate on the surface, effectively inhibit all but the most resistant plants, and are removed by wind to the adjacent plains. Some channels or posos seem to have been completely roofed over through such a process of progressive mineralization. Some seemingly undercut banks in the basin, and stone ridges, may have been produced by similar processes. Sinuous, elongate ridges of travertine-like materials that cross the basin floor may be attributed to lateral deposition of salts and roofing of channels, followed by recession of water levels, with subsequent deflation, compaction, and so on, leaving a "tube" exposed. Small structures of this type, formed similarly but through interactions of mosses, mineralized water, and high evaporative rates, are common near Monkey Spring, Arizona (Cole, 1968), and Blue Spring, southern New México (unpublished data).
Some travertine in the basin appears old, many deposits under water are alive in the sense of being actively formed by algal activities, and along many waterways, evaporitic travertines currently are forming. Periodic clearing of canals in the basin is necessitated by travertine deposition and by growth of aquatic plants. Many salts must also be bound in the flocculent, calcareous sediments. Extensive travertines now far removed from epigean waters evidence higher water tables, or additional springs, in the past. Detailed analyses of sediments from the area are badly needed to document past environments. A number of cores, ranging to 16 meters long, were taken in 1968, and are now under study. Hubbs, et al. (1965 ) reported one radiocarbon date of 2,070+/- 250 years before present, from peat samples from 2.25-2.35 meters in a core from Posos de la Becerra (SW1/4, I 12). Martin (in Hubbs and Miller, 1965) considered this as a "roughly median date" for springs of this type. Caution should be exercised in interpretation of radiocarbon dates from springs; Damon, et al. (1964) found anomalous dates of 17,000 to 25,000 years before present for living aquatic plants from Montezuma Well, Arizona. "Fossil" carbon dioxide from the spring water promoted this anomaly, and other such occurrences are known (Carl L. Hubbs, pers. comm.).
Despite processes of deposition described above, marshes persist in the Cuatro Ciénegas basin. In fact, they constitute the major aquatic habitat, comprising many square kilometers of the basin floor. Marshes are in association with lagunas, along streams, and also in isolated areas throughout the barrial. Most marsh habitats have clear, shallow water over soft bottoms ( Figs. 19 - 20 ). Only a few develop black and malodorous water in times of stagnation. Most receive a constant inflow, and in such areas sedges and other semi-aquatic plants become most luxuriant. Dominant species in the limited area studied by William S. Brown (unpublished) 4 were as follows, in order of decreasing abundance: Eleocharis rostellata, Scirpus olynei, and Spartina spartinae, Baccharis glutinosa, Cladium californicum, and Sporobolus airoides often form a conspicuous edge surrounding a marsh. Other plants recorded in similar habitats adjacent to rivers, canals, or lagunas, are Eleocharis macrostachya, Fimbristylis thermalis, Cyperus sp., Andropogon glomeratus, Phragmites communis, and Typha sp. The last occurs only in areas of relatively deep water. Large numbers of undetermined composites, plus other, less obvious plants, live near all suitable aquatic habitats.
{kind=link}
{kind=link}
Zonation of sedge species, as described by Bolen (1964) near salty waters in Utah, and by Cole and Whiteside (1965) for saline ponds in Arizona, has not been studied in the Cuatro Ciénegas basin; however, many of the communities are strikingly similar to those described by Bolen (Brown, unpublished).
Although most marshy areas of the basin are depressed below the adjacent land surface, some are obviously higher than the desert that surrounds them. Some are scarcely evident, but a number of the "raised marshes" are as high as a meter. They are totally water-filled; wet areas downslope are maintained by seepage from them. Meinzer and Kelton (1913) attributed similar hillocks in Pluvial Lake Cochise (Wilcox Playa, Arizona) to deflation; however, Hevly and Martin (1961) present data to support active formation of raised marshes adjacent to Croton Springs in the Wilcox Playa through accumulation of organic debris. Such deposits may provide valuable data on past conditions in desert basins, when studied.
The major factors presently affecting all kinds of marshes in the basin are periodic burning by local people, and canalization. The latter, of course, also seriously affects lagunas. Many posos and lagunas in the basin have water levels approaching the top of their stony banks. These are associated with extensive marshes, and have not been disturbed by canal construction. Canalization drops water levels locally, drains most marsh habitat, and leaves sloping, fine-grained soils as banks where travertine rims formerly occurred. Sedges and cattail, plus other plants, quickly colonize available soils, and a new riparian habitat is formed. Canalization does not totally destroy the laguna habitat because of the influx of water from deep aquifers, but marshes and shallow downflow habitats, plus most of the rock surfaces along banks, are removed from the aquatic system.
Barrial lakes, which were extensive prior to the construction of ditches and canals, are now represented in the basin by a few shallow depressions that hold permanent water. One such lake, Laguna Grande (SE1/4, J 11 , NE1/4, K 11; Figs. 4 and 18 ) has been studied most. It was for years considered an ephemeral playa with no inflow or outflow. In 1964, the outflow of Churince Laguna (center, K 12) was traced directly to the lake, and in 1965, it was confirmed that no epigean outflow exists. Laguna Grande is about a kilometer long, north to south, and perhaps half a kilometer wide. Depths do not exceed a meter. On its west and north sides are extensive dune-fields of white gypsum sand (Minckley and Cole, 1968a), which rise as high as io meters. Such morphometry appears typical of barrial lakes and playas in New México, Texas, and northern México (Parker and Whitfield, 1941; Germand, 1939; Judson, 1950; and others). On the southeast side is a now dry part of the lake, upon which porous "tufa" pyramids rise two meters above the floor. These structures are remnants of consolidated gypsum dunes rather than marl or travertine deposits as described from other desiccating basins (Jones, 1914; Dunn, 1953; and others). Ecology of this dying body of water presumably resembles that inferred from now-dry lakes of the American Southwest (Reed, 1930; Mitchill, 1956; Deevey, 1957; Cole, 1963, 1968); it is one of the few such lakes left in the region that retains faunal components other than brine shrimp (Artemia) or drought-resistant forms.
Environmental conditions in barrial lakes may be severe. Laguna Grande is not excessively mineralized, however, holding between 4 and 5 grams per liter of total dissolved solids. Cut-off pools along the shores often develop more than 400 grams per liter, and are essentially azoic. Bottoms in the lake are gray to white, relatively firm, and scarcely vegetated; they are covered in part by "algal biostromes" (Carozzi, 1962), some of which exceed 0.5 meter in diameter. Semiaquatic plants also are sparse. Temperatures fluctuate widely with insolation and evaporative cooling. Wind action and concomitant turbidities often are high.
Temporary waters of the Cuatro Ciénegas basin are diverse. Many formerly large barrial lakes now are ephemeral. Insofar as is known, most are too saline to support other than certain insects and phyllopods. Small depressions in the barrial, isolated from other waters and/or along the margins of other aquatic situations, usually develop fantastic concentrations of salts (Figs. 21 and 22 ). Borrow pits along newly constructed roads, tanks recently built along the mountains, and so on, are continually increasing the temporary habitat, and are yet to be studied.
{kind=link}
{kind=link}
Canals - Canal construction, especially in and regions, has greatly increased the available aquatic habitats (Pennak, 1958; Cole, 1963). In the Cuatro Ciénegas basin there are many old and new canals (Figs. 23 and 24 ) and construction proceeds apace. Some of the channels may have been built in the 16th Century, since Spanish-speaking peopleare known to have invaded the area at, or before, that time (Alessio Robles, 1938). There are no evidences that earlier, indigenous peoples practiced agriculture, or indeed, that they used aquatic habitats of the barrial at all, even for a water supply (W. W. Taylor, 1956, 1962, 1966), but semiaquatic plants were infrequently used in fabrication of mats (Brugge, unpublished; W. W. Taylor, pers. comm.).
{kind=link}
{kind=link}
Waters of the canals are generally cool, clear, and permanent except in periods of construction and repair. With three exceptions all canals have silt or marl bottoms at this writing. Canal de Anteojo (origin, SE1/4, E 12), was lined with concrete about 1950. Canal de la Polilla (origin, SE1/4', M 17) is being re-routed and lined, and Canal de Posos de la Becerra (origin, SW1/4,I 12 ) is being similarly constructed; concrete is soon to be applied to many canal banks. Dates of construction for the major canals of the area are given in Table 1, and were provided by José Lugo (pers. comm.), or are based on my observations.
The largest canal in the basin is the "New Canal," which bisects the barrial near its center (origin, NW1/4, G 17; Figs. 23 , and 27 ). This channel was built between 1958 and 1962, but remained relatively dry until 1963 when springs opened into its sides. It is yet to be connected to the Saca del Fuente Canal, but flows strongly at its lower end. In December 1965, discharge near Laguna de Ferriiio (N border, NW1/4, E 19) was estimated between 3 and 5 cubic meters per second. The canal is 8 toiLo meters wide at its top and perhaps five meters wide at the bottom. It ranges from 6 to 12 meters in over-all depth, depending on one's position along its length, with water depths of 2 to 4 meters. The canal is largely devoid of aquatic vegetation except in protected places where Typha and Potamogeton have established.
Most canals are smaller, resembling Canal de La Angostura (origin, SW1/4, F 12) which drains the Río Garabatal. This canal averages 1.5 meters wide and about 0.5 meters deep. Estimated discharges range from 0.1 to 0.5 cubic meters per second. Chara, Potamogeton, Naias guadalupensis, and Zannichellia palustris occur in its channel, but vegetation is periodically cleared by man. Thick stands of mesquite, numerous willows, some ash, and sparse sedges and other emergent plants line this, and other smaller canals ( Fig. 24 ).
SUMMARY OF THE AQUATIC FAUNA
Only a few groups of animals from the Cuatro Ciénegas basin have been studied, and the quality and quantity of information varies greatly from group to group. Birds (Urban, 1959 , W. K. Taylor, 1966) and mammals (Baker, 1956) have been generally studied in the State of Coahuila; probable lists of species in the basin can be determined from those works, and are not given here. Reptiles and amphibians are to be treated separately by Clarence J. McCoy, Jr. and Minckley (in preparation). Fishes are presently under intensive scrutiny, and have been since 1960. Invertebrates are scarcely known at all, especially the terrestrial groups. Stanley C. Williams (1968 and pers. comm.) has collected and studied scorpions, and he finds considerable endemism in the basin. Molluscs and crustaceans are becoming known, but existing collections have been made either by nonspecialists or by specialists in a short period of time and from limited habitats; other groups await study.
Many species remain undescribed, and though it seems ill-advised to list undescribed forms in a general summary, it is necessary in light of the ever-increasing study of the region by many people. I thank the numerous workers, acknowledged below, who have allowed announcement of their discoveries prior to independent publication.
Crustaceans - Seven kinds of aquatic crustaceans are known from permanent waters of the basin. One form, an eyeless, depigmented amphipod taken in 1968, is yet to be studied. Four are subterranean Isopoda, and are endemic in the area. Speocirolana thermydronis described by Cole and Minckley (1966) from a single specimen, but nowknown from additional material (Minckley and Cole, 1968b), is the most inland of the three species in that genus. Troglobitic cirolanids of the Western Hemisphere are presumably derived from marine forms that remained inland after recession of Cretaceous, or later, seas. With the exception of one species in the Appalachians, all are distributed around the margins of the Gulf of México (Bowman, 1964). In Cuatro Ciénegas, S. thermydronis lives in interstices of porous travertines at the margins of posos and lagunas, in sediments of spring runs, and in roots of riparian and aquatic plants, and is locally abundant.
The second genus of isopods, apparently an aberrant form of cirolanid, cannot be aligned with any described genus in North America. We found two species of this taxon in 1967 and 1968, along with S. thermydronis in travertine, bottom sediments, and fibrous bottom material or rubble. As with S. thermydronis, the new forms are white, eyeless, and highly secretive.
The fourth isopod is an asellote, also eyeless, that was taken after many hours of collecting. This form represents a new genus of the asellid subfamily Stenasellinae, an African-Eurasian group not before known in the Western Hemisphere (Cole and Minckley, 1969). Unlike the cirolanids, this species is blood-red in life, but is found in similar habitats, in springs and along the edge of lagunas.
The two other crustacean inhabitants of permanent waters both are described, and neither is endemic to the basin. The amphipod Hyalella azteca (Suassure) is exceedingly abundant in beds of aquatic vegetation, and beneath stones and rubble in spring sources and streams, throughout the area. H. azteca also is found within softer sediments of lagunas and posos, and rarely in temporary pools along other waters. This species enjoys a remarkably wide range in North America, and is highly variable in morphology; it is generally distributed in parts of northern México that I have visited. Palaemonetes suttkusi Smalley (1964), the seventh crustacean, is found in vegetated areas throughout the basin, and outside in the upper Río Salado de los Nadadores system; comparison of material from in- and outside detects no differences (in part, see Dennis F. Río, unpublished).6
Two other crustaceans have been taken in more temporary waters of the basin. One is an unidentified anostracan (fairy shrimp), represented in our collections by juveniles. The second is an harpacticoid copepod, Cletocamptus albuquerquensis (Herrick), which was caught in Laguna Salada, where it was abundant in water that held almost 400 grams per liter total dissolved solids (Minckley and Cole, 1968a). This species is widespread in highly saline waters (Wilson and Yeatman, 1959).
Molluscs - The following quotations from D. W. Taylor's ( 1966) summary of the Mollusca of the Cuatro Ciénegas basin suffice to review that fauna: "The small valley of Cuatro Ciénegas . . . has yielded the most spectacularly endemic fauna of freshwater snails known in the Western Hemisphere. The scanty collecting so far has been concentrated on only one habitat, the large springs, so that doubtless future additions will be made. Out of about 18 forms, only 13 can be identified specifically. One species is widespread in the region; the other 1 2 are new endemic species classified in five new endemic genera of which three represent new endemic subfamilies. This remarkably localized fauna shows scarcely any resemblance to known mollusks of eastern México, but the interior plateau of México is so poorly known that perhaps some groups will turn up elsewhere."
"Besides being strongly differentiated from the taxonomic standpoint, four of the new endemic genera are unusual morphologically. In having elaborate sculpture, color banding, a bristly surface, or being of large size for their groups they remind one more of marine snails than the groups to which they are anatomically similar."
"Out of five localized new genera, three are known only by shells from one locality each. The other three genera were collected alive at several localities, where they occur as isolated populations showing different degrees of morphological divergence. Two of the genera are not differentiated morphologically, but Mexipyrgus has undergone remarkable divergence. Seven populations sampled are assigned to six species. The morphological differences between these species are not correlated consistently with geographic location, so that they seem to have diverged through assortment of characters."
"No direct fossil evidence is available for judging the length of isolation of the endemic snails of the Cuatro Ciénegas area. If degree of taxonomic divergence is proportional to isolation, then the ancestry of some of the fauna reaches deep into the Tertiary, or Mesozoic. Mexipyrgus, an endemic genus of a widespread subfamily, shares characters of two genera, each having a fossil record; if all three genera diverged from a common ancestor they separated in early Tertiary times. By analogy with other species of Hydrobiidae the local endemic species of Mexipyrgus have evolved during the last 2 to 3 million years; if so, individual large springs or groups of springs have such antiquity."
"Two of the species that do not belong to endemic genera are nonetheless remarkable. Assiminea spec. is a rare inland occurrence of a usually coastal, marine family. Cochliopina milleri, spec. nov., is related to species of northern Guatemala rather than geographically nearby forms."
Additional collecting in the basin has revealed more species of aquatic molluscs, of which six are endemic. These, plus the species described by D. W. Taylor (1966), are listed in Table 2. One endemic, montane land snail also is known (Drake, 1951 ), and a number of other, disjunct population of terrestrial molluscs are known (Taylor, pers. comm.).
Fishes - The fish fauna of the Cuatro Ciénegas basin is unusually diverse for a desert area of its size. Nine families, 14 genera, and 20 species of fishes (and one possible introduction, yet unconfirmed) are present within or adjacent to the valley, and 9 of the 18 kinds now present inside the basin are endemic. Diversity of the fauna reflects the extent and variability of aquatic habitats, plus the influence of the relatively rich Río Salado (Rio Grande) fauna. By comparison, the extensive Bonneville system of Western United States has 5 families, 11 genera, and 21 species of fishes. The Lahontan complex supports 5, 10, and 13, respectively, and Death Valley has 3, 5, and l0 (Miller, 1958). Isolation of the last three systems is reflected, however, in a higher percentage of endemism of fishes. Sixty-two per cent of the Bonneville species are endemic, 69 per cent in Lahontan, and go per cent in Death Valley.
One of the most conspicuous fishes in the Cuatro Ciénegas basin is the Mexican tetra, Astyanax fasciatus mexicanus (Filippi). This is a mid-water fish, occupying almost all kinds of habitat with the possible exception of shallow marshes. It is abundant in deep, spring-fed lagunas where large, fast-moving schools range from the shoreline to as deep as lo meters, and in rivers and canals where it congregates below swifter areas. This aggressive characid is widely distributed in suitable habitats throughout northern México (Meek, 1904), where it is called "sardina," and is abundant outside the basin in the Río Salado de los Nadadores.
Three cyprinids live in the area. One is an undescribed endemic species of Notropis, which, like the tetra, lives in open waters of the larger habitats. The Notropis rarely schools with Astyanax, however, tending to travel nearer the bottom in groups of fewer than 15 individuals. In 1939, E. G. Marsh collected Notropis rutilus (Girard) within the basin at two localities south of Cuatro Ciénegas. The collections most likely came from La Angostura Canal ( Fig. 27), where Notropis sp. occurred before that segment of the canal was dried. No specimens of the endemic minnow were taken by Marsh, and N. rutilus now lives outside the basin in the Río Salado de los Nadadores. Reasons for this distributional change are indeterminable; other fishes taken by Marsh include basin endemics, negating the possibility for error in locality data. N. rutilus lives in the lower ends of turbulent riffles in the Río Salado, and often on deeper, strongly flowing riffles; neither of these habitats is now common in the basin itself.
The third cyprinid species in the fauna is Dionda episcopa Girard. Isolated populations of this fish occur through much of north México, restricted into disjunct populations through their proclivity for clear, spring-fed habitats that have little temperature variation (Hubbs and Brown, 1956). Morphological variation in D. episcopa is almost chaotic, making subspecific recognition of the numerous nominal forms presently unprofitable. Populations of D. episcopa in the isolated habitatsof Cuatro Ciénegas appear superficially uniform among themselves, but differ from the form in the Río Salado. When revisionary study is made the Cuatro Ciénegas form may be taxonomically recognized. D. episcopa lives near or on the bottom, generally in current rather than quiet pools. It is perhaps most abundant in canals, and enjoys a wide, nondistinctive distribution in the basin. All the cyprinids in- and outside the basin are called "sardinas," though residents recognize their distinctness from Astyanax.
Carp, Cyprinus carpio Linnaeus, are reportedly introduced in ponds (tanques) of the basin; however, none has been taken in the natural waters. Residents also report introduction of "carpas" into the Río Mesquites years ago.
No catostomids are known west of Puerto Sacramento; however, the gray redhorse ("matalote"), Moxostoma congestu albidum (Girard), occurs abundantly in the Río Salado de los Nadadores immediately to the east ( Fig. 2 ). Few young have been obtained, and those were near the lower ends of riffles. Adults concentrate in deep pools over silt or sand bottoms and near cover such as overhanging banks or thick beds of reeds (Phragmites). Similar habitats for M. c. congestum (Baird and Girard) in Texas were described by Hubbs, et al. ( 1953).
Two ictalurid catfishes inhabit the basin, and are also in the Río Salado. Both are cryptic and seldom caught unless especially hunted. A form of Ictalurus lupus Girard, the Mexican channel catfish, is widespread and common, living beneath cut banks and in dense beds of vegetation by day. At night, numbers of these jet-black fish feed slowly along the bottom, singly or in groups. This species also has been observed in underground channels, through which it must disperse with ease. The second species, Pilodictis olivaris (Rafinesque), is rare. Fewer than five specimens have been seen or collected. This fish is sought as food by local people, and is reported to reach 2.5 kilograms in weight. Fishermen angle in deep, strongly flowing streams, and near subterranean channels or deeply undercut banks, for this species. The channel catfish is locally known as "bagre," and the flathead catfish as "pildonte."
Gambusia marshi Minckley and Craddock is perhaps the most abundant fish in the Cuatro Ciénegas basin. This aggressive poeciliid ("lisa") is commonest in deeper marshes, marshy streams, and canals, and is least common in open channels and lagunas. In larger waters it sometimes swims in open water far from shore, foraging rapidly along the surface, but it is most concentrated along the banks of such habitats. Two color phases are present (Minckley, 1962) a dark phase is most common in the springs, and a spotted phase lives in canals, rivers, and more saline lakes and pools. No consistent morphological differences,other than color, have been found. However, Elena T. Arnold (unpublished) demonstrated marked mate selection by males of each color phase - homotypic females were selected almost invariably, and heterotypic females were rejected. Dark-phase fish have not been taken in the river outside the Cuatro Ciénegas basin, but spotted phase
G. marshi are abundant there.
A second species of mosquitofisb, Gambusia longispinis Minckley, is one of the basin's endemics and is unknown to local people. It is a small, secretive Gambusia, rarely living in water deeper than l0 centimeters, and almost never leaves dense beds of vegetation. In this habitat it occupies a mid-water or bottom position so that it and G. marshi are essentially allopatric, in the ecological sense, because of their respective habits.
Xiphophorus gordoni Miller and Minckley (1963), the third poeciliid, was described from, and is restricted to, the southeast lobe of the basin. There it occurs in three adjacent lagunas, and in a few small posos, and has habits and habitats similar to those of G. longispinis to which it is geographically allopatric. Kallman ( 1964) has studied histocompatibility in one population of X. gordoni using fin-transplantation techniques, and Jeanette H. Lysne (unpublished) detailed inter- and intrapopulation histocompatibility in three isolated stocks of this form. Rosen and Bailey (1963) synonymized X. gordoni with X. couchianus (Girard), but gave no reasons for their decision.
Another secretive species of the Cuatro Ciénegas basin, Lucania interioris Hubbs and Miller, inhabits small, environmentally extreme habitats. It is difficult to catch, however, and may be more widely distributed in larger habitats than is indicated at present. In marshes and saline pools the fish is rarely seen until seined. It dives into the substrate and remains buried except for the tip of its snout. Hubbs and Miller (1965) have discussed in detail the morphology, ecology, and distribution of this species.
Two additional cyprinodonts, both of the genus Cyprinodon, are endemic to the basin and have been the subject of intensive study since 1960. One of the species, Cyprinodon atrorus Miller 1968), ( has habits and habitats similar to most species of the genus (Miller, 1948), living in variable habitats of the basin floor (Minckley and Itzkowitz, 1967 ; Itzkowitz, unpublished). It is similar in its habitat selection and activity to L. interioris. The second form, Cyprinodon bifasciatus Miller, inhabits headwater springs and limnocrenes around the northern part of Sierra de San Marcos. In these, C. bifasciatus lives an open, active life that is quite dissimilar to that of other species. Young are on the bottom, foraging in schools that range from 10 to many hundred individuals. As they grow, they become less prone to contact the bottom; adults occupy open water, within a meter of the bottom, and range to the greatest depths of lagunas (deeper than lo meters in some). This fish has little regard for other fishes; even large predatory fish are occasionally chased by territorial males of C. bifasciatus. The two species of Cyprinodon are geographically sympatric in some parts of the basin, but are ecologically allopatric because of their divergent habitat preferences. Cyprinodon bifasciatus is highly sensitive to thermal changes, C. atrorus lives in highly variable habitats quite successfully.
Hybridization sometimes occurs between the two Cyprinodon. In undisturbed habitats, hybrids are infrequent and restricted to narrow zones of contact between the limnocrene or its outflow and marshes. Man's canalization has, however, extended the habitat available for C. bifasciatus, since water moving faster in canals than in natural channels retains its "limnocrene properties" far out into the basin floor. In summer, C. bifasciatus moves, or is carried, deep into the habitats of C. atrorus. This increases contact and allows mass hybridization. In smaller canals, hybrid swarms occur sporadically. In others, such as Canal de la Angostura, hybrid swarms in downflow areas far from parental influence stabilized morphologically, at least over seven years of study. The hybrid populations under study were destroyed in 1967 when a segment of the canal was abandoned and dried. The report of this phenomenal system of hybridization is in preparation (Minckley, unpublished data).
Sunfishes comprise a minor numerical component of the biomass in the basin, but one, the largemouth bass, Micropterus salmoides (Lacépede), is the omnipresent "top carnivore" in open waters of all the larger habitats. In the central part of northern México this fish is called "robalo," a name applied to the marine snook (Centropomus) in coastal areas. The second species of sunfish is best referred to Lepomis megalotis (Rafinesque), the long-ear sunfish. This species, locally "moiarra rojo" because of the brilliant red sides of breeding males, is uncommon. It lives near ledges and other cover in largersprings, lakes, and streams. Substantial morphological differences are apparent when specimens of both centrarchid species from the basin are compared with fish from the Río Salado. However, since no data are presently available, the magnitude of differentiation is not definable.
One darter of the genus Etheostoma is present, but rare, in the basin. It has been caught from the Río Mesquites and some closely associated drainages due south of Cuatro Ciénegas. This fish shares many characters with Etheostonw grahami (Girard) that inhabits the Río Grande drainage, and with E. lepidum (Baird and Girard), an endemic in streams of the Edwards Plateau of Texas (Hubbs, 1954; Moore, 1968). In addition, substantial similarities exist between the basin form and E. pottsi (Girard), which lives in México to the south and west of Cuatro Ciénegas (Meek, 1904). Revisionary study is needed to adequately interpret the darters of the Río Grande and southern drainages in México.
Cichlids present one of the most challenging situations yet found in the basin. Four kinds are present: three undescribed species of the genus Cichlasoma, and the common Río Grande cichlid, Cichlmoma cyanoguttatum (Baird and Girard). The preliminary announcement of this problem by Taylor and Minckley (1966) will suffice for the present account. "These three [endemic cichlids] differ from each other and from the Río Grande cichlid in their feeding adaptations, and parallel the striking modifications that cichlids have undergone in the ancient African Great Lakes. . . . One cichlid has thick, heavy teeth in the throat region, used for crushing the snails that are its main food. Snails are not eaten by the other species. A second form is almost identical externally, but has very fine teeth, lacks the strong crushing muscles, and feeds mostly on bottom debris, or other soft organic matter. A third form is recognizably different in shape. Its elongate form, strong jaws and associated structures are correlated with a diet made up principally of other fishes. The localized occurrence of these three species, the unusual specialization they show in food and feeding, and the general lack of external differences suggest that they evolved within the Cuatro Ciénegas area. They are probably adaptations to adjacent nicbes, for isolation in separate systems would not have favored specialization in diet."
In distribution, C. cyanoguttatum is generally on the east side of the basin, as far west as El Mojarral (SE1/4, G 12, SW1/4, G 13). The endemic forms broadly overlap C. cyanoguttatum in range; they occupy most waters west of Laguna Tío Quintero (SW1/4, NE1/4, L 16). The "finetoothed form" is most widespread of the endemic species, living in lagunas and in rivers and canals. The other two kinds are generallyrestricted to the larger limnocrenes and streams. All the forms appear similar in habits-, with young living with the female for a while, then moving to shallows, and as adults occupying open water or residing near various kinds of cover.
GEOLOGIC BACKGROUND
Structural Features and Their History
The antiquity of the Cuatro Ciénegas basin, as indicated by its unique, relicted fauna, is borne out by data available on the geology of the Mesa del Norte (Murray, 1961 ; Weidie and Murray, 1967)Predominately Mesozoic strata of the present mountains of Coahuila rest on a basement of marine Permian sediment and some post-Permian intrusives to the south (Böse, 1921 ; Mullerried, 1942; Kellum, et al., 1936). Permian sedimentary deposits probably were formed in an extension of the West Texas Geosyncline of that age (Böse, 1923; King, 1934). In central Coahuila, the basement is composed of granitic formations, and in the north, pre-Cambrian strata outcrop in Sierra del Carmen (Baker, in Böse, 1923). The last formation, and Paleozoic rocks of the Potrero de la Mula just north of Ocampo reported by Kellum, et al. (1936; see, however, Murray, 1961 ), may represent parts of an ancient land mass that existed adjacent to the western Permian sea.
Paleozoic surfaces were truncated in early Mesozoic when the Coahuila Mountains, an extension southward of the Marathon-Ouachita structural axis, were uplifted (Maldonado-Koerdell, 1964). The sea transgressed into the complex "Mexican Geosyncline" (see Murray, 1961) that existed across much of north-central México from late Triassic through Jurassic and early Cretaceous (Maldonado-Koerdell, 1958, 1964). Stratigraphic variations in sediments deposited in this shallow sea, in the area east and south of the present site of Cuatro Ciénegas, indicate a fluctuant shoreline along the southern edge of a persistent block, called the Coahuilan Peninsula (Burkhardt, 1906, 1930; Imlay, 1936; Kellum, 1932, 1936; Kelley, 1936; Murray, 1961; and others). Mid- and early Cretaceous seas covered this peninsula, inundating all of northern Coahuila by Comanchean times (Böse and Cavins, 1927; Weidie and Murray, 1967). Facies of gypsum in the central and eastern parts of the state (Imlay, 1936; Kellum, 1936; Jones, 1938; Murray, 1961 ) indicate sedimentation in isolated shoreline lagoons formed during slow encroachment and recession of seas over the persistent foreland (Kellum, et al., 1936). Interbedding oflimestones and clastic materials with gypsum further substantiates longand short-term oscillations of sea level during this period.
The Laramide Revolution of early Upper Cretaceous initiated a progressive regression of seas from geosynclinal areas. Early stages of orogeny, in west and central México and in the northern part of the geosyncline (including south Texas and northern Coahuila), were accompanied by subsidence in the Coahuilan Trough that now includes the area of Laguna de MayrAn. Subsidence also occurred in an extension of the trough along the east side of the old Coahuilan Peninsula (Imlay,1936; Kellum, et al., 1936; Arellano, 1951); this structure is called the Sabinas Basin by Murray (1961). At close of Cretaceous, seas withdrew -and the northern part of the area was elevated with initial phases of structural deformation; the Coahuilan Trough remained inundated by shallow seas until early Tertiary time (Weidie and Murray, 1967).
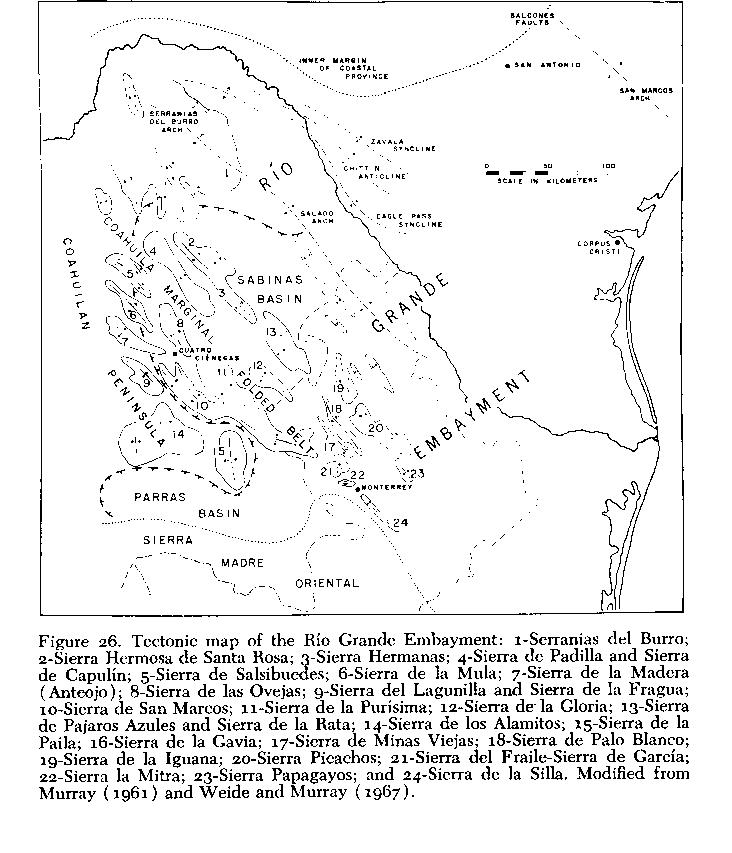
The Tertiary stage was therefore set for severe deformation and structural modifications in the area, which, along with erosion (West, 1964), produced the land surfaces seen today ( Fig. 26 ). Marine tradsgressions in Oligocene (Maldonado-Koerdell, 1964) did not reach the area of Cuatro Ciénegas. The geosynclinal area was folded in late Paleocene and in Eocene, along with some faulting and vulcanism (Kellum, et al., 1936; Imlay, 1936; Maldonado-Koerdell, 1964; and others). Next followed contraction, providing a phase of faulting in mid Tertiary, then dominantly vertical uplift that perhaps continued with decreasing intensity into Quaternary (Weidie and Murray, 1967). This produced in its later stages a topography like that of the Basin and Range Province in western United States (Thayer, 1916; Robles Ramos, 1942).
{kind=link}
During Tertiary, pressures from the west, south, and east (Kellum, et al., 1936) deformed upper Cretaceous strata overlying and surrounding the foreland of the Coahuilan Peninsula. These processes are indicated by strata overturned toward the northeast in the Sierra de Mapimi (Singewald, 1936) and in other mountains of the area west of the "Laguna District" near Torreón, Coahuila (Kellum, 1936), by north-facing overthrusts and recumbent folds in the Sierra de Parras (Imlay, 1936), and by northwest to west-southwest overturns in folds of the eastern side (Kellum, et al., 1936). Cuatro Ciénegas is near the eastern margin of the old Coahuilan Peninsula, in an area that may have been relatively unaffected by these orogenic forces (Weidie and Murray, 1967).
There is little evidence for Recent crustal movements in the basin, except that the entirety of the north end of Sierra de San Marcos, when compared with the extreme dissection of its south end (Kellum, et al., 1936), may indicate progressive uplift of that mountain. Major structural features of Coahuila, as noted above, are believed to have formed concomitant with the Laramide Revolution in late Cretaceous and the first part of Tertiary. Keith (1928) believed that the Laramide continued into Miocene, and that normal faulting began in Miocene and extended to Recent, at least locally. Upwarping, with associated minor diastropbism, seems to have prevailed in northern México through Quaternary (Kellum, et al., 1936; Weidie and Murray, 1967). There is little evidence for major orogenic disturbances in northern Coahuila in laterPliocene and early Pleistocene as was stressed by Schuchert ( 1935) for the Mexican Plateau in general. Igneous extrusives of late Tertiary times are tilted, and are severely eroded, but are not folded or otherwise modified (Singewald, 1936; Kellum, et al., 1936). The same appears true for some lacustrine sediments deposited in the Coahuilan Trough (Kelly, 1936; Weidie and Murray, 1967). The mid- and late Tertiary conglomerates deposited in intermontane basins and east of the mountains on the Coahuilan Piedmont (Porter, 1932; Imlay, 1.936), likewise show little deformation.
Drainage Relations
Regional - Erosional and drainage data outlined for the Valle de Sobaco area (Kelly, 1936; Imlay, 1936), apply to the Cuatro Ciénegas region. First, certain canyons in Sierra de Parras, including those occupied by the Ríos Nazas and Aguanaval, appear to represent drainages that originated on an ancient land surface that was approaching base-level. These valleys incised with uplift, and were deepened in a succeeding erosional cycle, cutting across anticlinal ridges and other structures. Boulder conglomerates pass laterally into chalky limestones, indicating that internal drainage may have existed locally at this early stage, in a region of high topographic relief (Kellum, 1936). Valley cuts superimposed on Sierra de la Fragua, west of the Cuatro Ciénegas basin, the Cañon del Agua, and the puertos to the east of Cuatro Ciénegas, appear also to be part of antecedent system(s). Most valleys were presumably desiccated by uplift that exceeded erosional capabilities of their through-flowing streams; formation of extensive alluvial fans was induced in many places.
A wetter stage resulted in alluviation of lower canyons and developmedt of extensive fills. Marls interbed with gravels in many areas, and in later stages increasing aridity is indicated. This phase included a period of intensive erosion of plains along the mountain fronts. Some stream channels were left banging and others cut deeply into conglomerate deposits of the preceding period. In the most recent stages, everincreasing aridity is indicated by erosional features and notably by lacustrine deposits left high above existing valley floors. Some of these deposits may, in some cases, have been elevated by tectonic movement, but the horizontal position of most diminishes this probability. Consolidated conglomerates along Río Cañon, in Cañondel Agua north of Cuatro Ciénegas, and in puertos leading east from the basin ( Fig. 2), are made up primarily of angular talus debris. These, and a few other formations along the mountains (especially near Rancho San Marcos; NE1/4, SE1/4, L 11), appear to represent erosional remnants of (Tertiary?) land surfaces similar to those described farther south.
Much evidence points toward the past presence of large lakes in the Cuatro Ciénegas basin. However, field inspection of the margins of the basin reveal no visible terraces, nor beach deposits, to document such high-level Pluvial (or pre-Pluvial) habitats. Only one small butte just south of Rancho San Marcos may be a remnant of lacustrine deposition. It is of travertine, about 30 to 40 meters high, and is slightly tilted toward Sierra de San Marcos ( Fig. 25 ). This structure is not mentioned in geologic literature available to me. In fact, the works of Kellum, et al., (1936), and others cited therein, mention no major lacustrine deposits in northern Coahuila.
{kind=link}
It is not surprising that terraces and beach lines are absent near Cuatro Ciénegas. In most larger desert basins, and possibly in all such basins of Western North America south of the Great Basin that depend on their immediate drainage for water, such marks may never have been formed. Pleistocene Lake Otero, New México, laid more than 70 meters of sediment in its center (Herrick, 1904), but never rose high enough on its walls for a long enough period to cut now-recognizable shoreline features (Kottlowski, 1958). Basins with outlets relatively high above their floors cut extensive terraces because of stabilization at outlet level. This is exemplified by the Provo Shoreline of Pluvial Lake Bonneville (Morrison, 1965), formed at outflow level of the Provo Standstill. The lack of high lake levels in southern basins, and especially in Cuatro Ciénegas, is attributable to persistent drainage through deep, antecedent channels that breach the surrounding mountains.
Isolation of the Cuatro Ciénegas basin was almost certainly intermittent. Closure was periodically effected by desiccation, uplift of anticlinal structures, by accompanying warping of the Plateau in general (Kellum, et al., 1936), and by breakdown in its western part. Arellano (1951) theorized that uplift in the southern (or western) part of the Bolsón of Coahuila toward the east or north, early in Pleistocene, spilled ancestral Lago de Mayrán. Uplift in the eastern Sierra Madre Oriental reversed this trend later in Pleistocene, and again closed that basin. It is probable that similar uplift on the south and west sides of the Cuatro Ciénegas basin contributed to its connections (or spillage) into the Río Grande basin. Short periods of isolation, in the geologic sense of time, are very important from the standpoint of biologic differentiation (Hubbs and Miller, 1948), and may have been induced in the Cuatro Ciénegas area through periodic dune formation, alluviation of puertos, or by mineral deposition in lower segments of watercourses. There can be little doubt that in more or less closed basins with in-flowing rivers (e.g., Bolsón de Coahuila), or with large springs such as those near Cuatro Ciénegas, increased rainfall that occurred periodically in the past produced major lacustrine environments. The extent and influence of these habitats on the history of the biota remains largely undetermined.
Existing structural data render untenable the suggestion by Conant (1963) that the more southern basins of the Ríos Nazas and Aguanaval drained northeast toward, and perhaps through, the Cuatro Ciénegas area, by filling and overflowing a series of Pluvial lakes. Such an integrative sequence for the present Río Grande system was postulated by King (1935), as opposed to antecedence. In the Mesa del Norte of México the tendency for disruption of the north-south-trending axes of the mountain ranges by the Sierra de la Madera (equals Sierra de Anteojo), and by structures south-southwest of that range ( Fig. 26 ), might have allowed an inflow of drainage from the Torreón area to the northeast, but the tendency for orientation of ranges from east to west in the area west and south of, but not including Sierra de San Marcos, would weaken this possibility. Breaching of the east-west axis seems most probable far west of the Cuatro Ciénegas basin. Incision of the north-south axes appears less likely, anywhere but to the east toward Saltillo (Arellano, 1951), than is a pathway, suggested by Meek (1904) along the route now marked by the political boundary between the states of Coahuila and Chihuahua.
Intra-basin Drainages -Data from topographic maps and aerial photographs are now available from which to define the historic drainages of the Cuatro Ciénegas basin. However, underground connections occur between many parts of the aquatic system; with continuing discovery of these channels, and the rapid, natural colonizations of newly foundered posos by fishes, I am hesitant to consider any of the drainages as totally distinct. Present divides between adjacent parts of these systems are in places only a few meters high and a few meters broad. Subterranean exchanges of water obviously occur, and, with slight increases in water levels, surface flow would certainly become established. In the following discussion these reservations should be kept in mind.
Some drainages at the time of Spanish invasion may be inferred from early maps; however, reliability of some of these is questionable. For example, a fragment of the relatively late "Carta General del Imperio Mexicano," dated 1860, shows two huge lakes south of Cuatro Ciénegas (Alessio Robles, 1938). These are called Lago de Santa Maria and Lago de Agua Verde. The former may now be represented by Laguna Grande, and the latter by the now-dry sump(s) on the east side of thebasin. Alexander von Humbolt's 1811 map (in Alessio Robles, 1938) shows Lago de Agua Verde as the origin of the Río Salado de los Nadadores. Aerial photographs and ground surveys support the presence of at least two major barrial lakes, and in the general vicinity of the two pictured in the 1860 map. However, the name Agua Verde now is applied to a place near Ocampo, north of Cuatro Ciénegas ( Fig. 1).< p> An unpublished 1887 map of landholdings in the Cuatro Ciénegas basin indicates that the Río de San Marcos (equals Río Mesquites) originated Dear what now is Rancho Tierra Blanca (N border, NW1/4, H 12) and terminated in a large lake on the east side. Confirmation of such a drainage pattern by Rodriguez Gonzáles (1926) was mentioned before. If no outflow occurred prior to canalization, other than that from the laguna on the northeast side (D 20), and Río Polilla (origin, SE1/4, F 21), the collective discharge of Río Puente Chiquito, Río Mesquites, and other lesser waters, must have resulted in a considerable lake(s).
There are seven major epigean drainages in the Cuatro Ciénegas basin, perhaps persisting since Pluvial times, but surely connected, in part at least, when greater volumes of water were present. These differ from those listed by Hubbs and Miller (1965).
I)The now-isolated system originating in Laguna Churince (center, K 12), and terminating in Laguna Grande (SE1/4, J 11, NE1/4, K 11), including all the area to the south (Laguna de San Marcos, NE1/4, SE1/4, L 11; dried in 1965-6), is the least disturbed drainage in the basin. It is presently isolated from the following system by a low travertine sill and by dunes.
II)A major system, originating in Posos de la Becerra (SW1/4, I 12) and flowing west of the complex of rivers and lagunas called El Garabatal (N1/2, H 11), now feeds the canals de la Becerra and La Angostura. Prior to alteration this system passed west and north of the Río Mesquites system (described next) to enter what now is a playa south of Cuatro Ciénegas (F-G, 13-16; not outlined on Fig. 27 from which it possibly overflowed into Río Mesquites and/or Río Puente Chiquito (IV, below) in times of high water levels.
III)Rio Mesquites, described before (PP. 13-15), is the largest integrated system in the basin. Much of its water originates at Tierra Blanca (N border, NW1/4, H 12), a thermal rheocrene (31' Centigrade), but it receives cool spring inflows (22' to .23.5' Centigrade) that originate at El Garabatal (N1/2, H 11) suspiciously near the river from la Becerra. It seems likely that underflow from the Becerra channel (Rio Garabatal) is here represented, but no direct surface connection has been discovered in the field or is discernable on aerial photographs. Río Mesquites derives additional complex inflow along its course, then is diverted into Canal de Saca del Fuente (origin, S,1/2, NW1/4, H 15). Before diversion it passed east through a series of marshes and lagunas, to sump in an eastern barrial lake (E-H, 17-20; not outlined on Fig. 27 ). "Lagunas de los Burros (SE1/4, NW1/4, H 16) and Mesquites (NW1/4, H 17) persist, with meager inflow, in the old river channel east of Canal de Saca del Fuente.
IV)Rio Puente Chiquito (main origin, NW1/4, NW,1/4, H 15), also discussed earlier (P. 15), is fed by cool waters throughout its system (21' to 24' Centigrade). All spring origins of this river that have been visited emerge from large travertine-lined tubes. As with Río Mesquites and la Becerra channels, Río Puente Chiquito and Río Mesquites are closely approximated, especially in the area of Canal de Saca del Fuente ( Fig. 27 ). Ground survey in 1968 revealed probable connections in this area, in periods of high water, but no natural surface flow was present. Río Puente Chiquito , prior to Canal de Saca del Fuente, flowed at least partially into the same barrial lake as Río Mesquites. It may also have been connected directly to the Río Grande system via a low area along the north side of the basin, and thence through channels near Laguna de Ferriño (N border, NW1/4, E 19).
V)A fifth system originates principally in Laguna Tío Candido (NW1/4, NE1/4, J 14) and (originally) in a large, thermal spring, Escobeda (E1/2, NW1/4, I 14), that is now canalized into the Río Mesquites. Water from Tío Candido passes into marshes, and underground, to emerge at Lagunas de los Hundidos (N1/2, J 16); subsurface flow is resumed, and water reappears near Lagunas de San Pablo (NE1/4, I 17). The subterranean channel(s) is obvious on aerial photographs, well marked by vegetational change, and in the field by slight depression of the land surface and the presence of sinkholes. Lagunas de San Pablo connects, naturally through marshes and artificially through a small canal, to a complex of lagunas (H 19, unnamed) at the south end of the barrial lake (now a playa) on the east side.
VI)The Santa Tecla system, now drained by Canal de la Polilla (origin in Laguna de los Fresnos, SE1/4, M 17), is largely disjunct, and the southeast lobe in which it is located ( Fig. 27 ) has been relatively inaccessible until 1967. A remarkable number of posos are present in this system (K 16 , L 16-17, M 17 ); many of these have no surficial connections. This complex, on the basis of interpretation of aerial photographs, drained north-northeast to enter the south end of thehuge eastern barrial lake. The organic sediments downflow north in this system indicate an extensive marsh in the past that was generally devoid of massive salt deposition. Total dissolved solids in waters from this system are far lower than in other waters of the basin (Minckley and Cole, 1968a).
VII)The seventh major system, the Río Salado de los Nadadores (Rio Grande), headed in D20 on the north, and near the eastern mountain to the south (F 21). No natural channels of this system now carry permanent water. Isolated waters occur that have no apparent connections. Only extensive field work can clarify their relations to other waters of the area.
DISCUSSION
No other place in North America offers greater opportunity for the study of aquatic and semiaquatic systems in deserts than does the Cuatro Ciénegas basin. It is located in a zone of extreme aridity, yet the persistence of thermal springs presents an almost-unlimited spectrum of aquatic and mesic habitats. The rich, varied, and highly endemic biota increases the information content even more. Most importantly, all this diversity is in a small, accessible area, that can be successfully toured by modern vehicles in a day.
A major purpose of this discussion is to emphasize the urgent need to study, salvage, and protect this and other such areas, in deserts. Spring-fed pools and associated habitats are rapidly disappearing as man invades and occupies and lands (Miller, 1961: Minckley and Deacon, 1968). Each isolated spring, or spring system, unless too hot, saline, or otherwise inhospitable, contains biotic elements that represent either remnants of formerly widespread forms, or special endemics. Many such habitats are destroyed, especially in Southwestern United States, and a number of known Mexican localities are highly modified, drying, or dried. Such extinction of isolated water bodies is more serious than drying of a stream tributary to a larger, integrated system. The latter may extirpate a large number of organisms, reducing the amount of habitat available for a species' use. However, the former kind of catastrophe may eliminate the entire gene pool of a species, at one stroke. Increasing technological application in the Cuatro Ciénegas region, as elsewhere, is accelerating changes, and the extinction of some springs seems imminent.
A pattern emerging from studies of the Cuatro Ciénegas biota implies considerable antiquity and continuity through time of the aquatic habitats. Relationships differ markedly from group to group, yet someof the species in each taxon studied are highly differentiated from their living relatives and must date from extreme antiquity. This is especially evident in the molluscs (D. W. Taylor, 1966, and unpublished), and perhaps in the crustaceans (Cole and Minckley, 1966, ig6g, and unpublished). On the other hand, some organisms are weakly differentiated from adjacent populations, or not changed at all; 5o per cent of the fishes occur as similar morphotypes in- and outside the basin, and the more vagile species of invertebrates also fall in this category. Criteria based on "degrees of difference" in all the groups presently produce a mosaic of indicated differentiation. Multiple invasions of the area, with differing time regimes resulting in varied differentiation, are clearly indicated. Geologic stability in the Cuatro Ciénegas area may account for persistence of ancient faunal elements. Changes in Pleistocene (reviewed for areas north and south of Cuatro Ciénegas by Martin and Harrell, 1957; Martin, 1958a-b; and Martin and Mehringer, 1965), as inferred from studies elsewhere, may be used to account for some more of the complex fauna, and changes produced by
man can explain recent faunal additions.
Much actual work is needed in the basin to support existing inferences, and acceleration of modification by man adds some urgency to the situation. The biota is definitely under stress, and even the deposits associated with springs and the extensive dunes of the barrial are soon to be commercially exploited. The answers needed to interpret past conditions may exist in the deposits, and this aspect needs rapid attention.
Papers on the basin and its biota, including this one, are mostly descriptive. Synthetic work, however, will soon be underway. Diversity of the biota, coupled with simplified habitats of constant-condition springs, makes comparative ecological research highly rewarding. Few places other than desert springs have a combination of constant input, high and relatively invariable insolation, and varied habitat. Information potential of the area is unlimited, research has just begun.
GAZETTEER OF LOCALITIES
Place names may refer to areas rather than to specific points on a map; some are specific, however, and are marked with an asterisk (*) in the following list. Spelling follows local usage. Where more than one name is applied, the one least frequently used is in parentheses. Names coined by us are given in quotations. Coded locality data refer to the grid provided in Figure 27 , letters (D-M) are from north to south and numerals (11-23) are from west to east. Distances given in kilometers are straight-line measurements from the center of Cuatro Ciénegas. See Figures 1 , 2 , and 26 for additional geographic information.
Lagunas, Laguna Systems, and Posos (Pozos)
*Anteojo (Laguna Anteojo), S1/4, SE1/4 , E 12
*"Laguna de los Burros, SE1/4 NW1/4, H 16.
Chiqueros (Lagunas de Chiqueros), SW1/4, NE1/4, I 11
*Churince (Laguna Churince, Posos Bonitos), Center, K 12
*Escobeda (Escobedo), E1/2, NW1/4, I 14.
*Laguna de Ferriño, N border, NW1/4, E 19 (dried in 1966-7).
Los Fresnos (Lagunas de los Fresnos),SE1/4, M 17.
El Garabatal (Lagunas del Garabatal), western tributaries of Río Mesquites (not Río GarabataI, see below), N1/2, H 11
*Laguna Grande (Lago de Churince), SE1/4 , J 11 , and NE1/4, K 11.
Los Hundidos (Altamira, Lagunas de los Hundidos), N1/2, J 16.
Juan Santos (Lagunas de Juan Santos), Center, W border, H12. "Laguna Mesquites," NW1/4 , H 17.
El Mojarral, SE1/4 , G12, and SW,14', G'3 (two major lagunas named "west" and "east").
*Laguna Orozco, NW1/4, NE1/4, J 14.
*Posos de la Becerra, SW1/4, I 12.
*Laguna Salada, SW1/4 , SW1/4, F 14 (frequently dry, filled sporadically for evaporative collection of salts).
*San Marcos (Laguna de San Marcos), NE1/4, SE1/4, L 11 (dried in 1965-6).
San Pablo (Lagunas de San Pablo), NE1/4, I 17.
*Laguna Santa Tecla (Santa Tecla), SW1/4, SW1/4, M 18.
*tio Candido (Laguna Tío Candido), NW,4, NE1/4, J 14.
*Quintero (tio Quintero), SW1/4, NE1/4, L 16.
Rivers and Creeks, Including Place Names for
Localities along the Channels -
*Rio Cañon, origin ca. 10 km NNW of Cuatro Ciénegas (north of D 13); flow through Cañon del Agua (north of D 13-14); termination in domestic water supply and irrigation, Cuatro Ciénegas (E 15).
*Rio Churince, origin in Laguna Churince; termination in Laguna Grande (see above).
*Rio Garabatal, origin in Posos de la Becerra (not in El Garabatal, see above termination in Canal de La Angostura,S W1/4, F 12.
*Rio Mesquites, origins in E2/3, N1/2, H 11, and NW1/4, NW1/4, H 12; termination in Canal de Saca del Fuente, N border, SW' 4, H 15. Tierra Blanca (thermal spring), N border, NW1/4, HIL2. Puente San Marcos, NW1/4, SE1/4, G 12. Los Corrales (meandering, wide portion of river), principally H 13. Puente Orosco (Los Corrientes), N border, SE1/4 H 14. Dos Puentes, at divers;on of river into Canal de Saca del Fuente, S1/2, NW1/4, H 15.
*Rio Positos (Los Positos), tiny stream at NE1/4, F 17; almost dry in 1960, not again visited.
*Rio Polilla, origin SE1/4, F 21; flowed out of basin at Puerto Salado, D 23 (long dry). 'Rio Puente Chiquito, origins S1/2, G 14, and N1/2, H 15; termination in Canal de Saca del Fuente, El/2, SW1/4, G 16. Puente Chiquito, S1/2, SE1/4, G 15.
*Rio Puente Colorado, origin NW1/4, SE1/4, H 14; termination in Río Mesquites in times of high water near Puente Orosco (see above); now receives Canal de Escobeda. Puente Colorado (now impassable), just below inflow of Canal de Escobeda.
Canals (See also Table 1)
*Anteojo, origin in Laguna Anteojo, see above; termination in fields, E 15-16.
*La Angostura, origin in Río Garabatal, SW1/4, F 12; termination in fields, E 16-18 (may connect, occasionally, with Canal de Saca del Fuente in E 18).
'Escobeda, origin in Laguna Escobeda; terminates in Río Puente Colorado, see above.
*Ferriño, origin in northern tributary of Río Mesquites, NE1/4, SW1/4, G 12; termination in fields, E 15-16.
'Julio, origin in northern tributary of Río Puente Chiquito, SW1/4, G 15; termination in fields, E-F, 16.
"'New Canal," origin NW1/4, G 17; flows Out of basin at Puerto Salado, D 23.
'Orosco, origin in Tío Candido; terminates in Río Mesquites at Dos Puentes, see above.
'Polilla (Rio Polilla), origin in Lagunas de los Fresnos, see above; flows out of basin at Puerto Salado, D 23.
'Saca del Fuente, origin in Río Mesquites at Dos Puentes, see above; flows out of basin at Puerto Salado, D 23.
'Posos de la Becerra, origin in Posos de la Becerra, see above; not completed in 1968 (to flow out of basin at Puerto Salado, D 23).
Towns and Ranches (See also Figs. 1 - 2 )
*Boquillas, SW1/4, NE1/4, D 21.
*Rancho Orozco, NW1/2, NE1/4, L 14.
*San Juan, SE1/4, N1/4, D 21.
*Rancho San Marcos, NE1/4, SE1/4, L11.
*Rancho San Pablo, NW1/4, L 21.
*Rancho La Vega, W1/2, J 22.
*Ejido Santa Tecla, W 1/2, SE1/4, M 17.
*Rancho Tierra Blanca, N border, NW1/4, H 12.
Early work in the Cuatro Ciénegas basin was supported by a grant-in-aid from Sigma Xi-RESA funds, and by a faculty research grant from Western Michigan University (1961-63). Later studies have been funded by National Science Foundation Grants GB-2461 and GB-6477X.
John M. Legler first allowed me an opportunity to visit the Cuatro Ciénegas basin in 1958. In 1960, J. E. Craddock, C. B. Stone, and D. R. Tindall, pooled their "vacation funds" with mine, and in a vehicle provided by the University of Louisville, we made the first detailed collections from the area. C. L. Hubbs and R. R. Miller then extended assistance, and I accompanied them to the basin in 1961. Since then, more than thirty persons have contributed to field work in the area. A. T. Hotchkiss, D. J. Pinkava, and D. R. Tindall identified most of the plants and have done independent work on the flora. Work on molluscs by D. W. Taylor, and on pbysico-chemical limnology and crustaceans by G. A. Cole, have broadened the over-all study and uncovered many new avenues for future explorations. S. C. Williams is now studying the scorpions, and C. J. McCoy, Jr. is dealing with reptiles and amphibians. The last four persons allowed me to consult their unpublished data in preparing this report. The following also deserve special thanks for their help and interest: E. T. Arnold, G. A. Batchelder, W. E. Barber, W. G. Bradley and students, W. S. Brown, A. C. Echternacbt, M. Itzkowitz, D. W. Johnson, J. E. Johnson, B. Knowles, R. K. and J. Koehn, J. J. Landye, P. S. and J. H. Lysne, G. Lytle, C. 0. Minckley, W. S. Parker, J. N. Rinne, W. K. Taylor, A. P. Wick, and D. C. Williams.
G. A. Cole, C. L. Hubbs, R. R. Miller, and D. W. Taylor read this report and offered helpful and constructive criticisms. All photographs from 1960 were taken by C. B. Stone; other photographs are by myself, G. A. Cole, or commercial photographers.
Officials in México have cooperated fully in providing permits and letters of introduction to expedite research in northern Coahuila. It is hoped that this paper will repay somewhat for their patient assistance. Daniel Rodriguez Villarreal, Modesto de la Garza Palos, José lbarra Gonzáles, and Francisco Manrique DAvila are a few of the people in Cuatro Ciénegas who have provided much needed assistance. Without the help of José Lugo Guajardo and his wife Catalina Diaz de Lugo, very few of the data given here would even have been gathered. "Pepe" Lugo and his family deserve special thanks from all people associated with Cuatro Ciénegas studies.
Last, I acknowledge the patience and assistance provided by my wife, Barbara, and my family, both at home and in México.
LITERATURE CITED
Alessio Robles, V.
1938. Coahuila y Texas en la Epoca Colonial. Editorial Cultura, México, D. F.
Arellano, A. R. V.
1951. Research on the continental neogene of México. Amer. J. Sci., 249: 604-616.
Baker, R. H.
1956. Mammals of Coahuila, México. Univ. Kans. Publ., Mus. Nat. Hist., 9: 125-335.
Boke, N. H.
1968. Excursiones de la Universidad de Oklahoma en el Norte de México - 1967. Cactaceaes y Suculentas Mexicanas, 13: 50-58.
Bolen, E. G.
1964. Plant ecology of spring-fed salt marshes in western Utah. Ecol. Monogr., 34: 3.43-166.
Böse, E.
1921. On the Permian of Coahuila, northern México. Amer. J. Sci., Ser. 5, 1: 187-194.
1923. Vestiges of an ancient continent in northeast México. Ibid. 6: .127-136, 196- 214, 310-337.
Böse, E., and 0. A. Cavins
1927. The Cretaceous and Tertiary of southern Texas and northern México. Univ. Tex. Bull., 2748: 7-142, 1 pl.
Bowman, T. E.
1964. Antrolana lira, a new genus and species of troglobitic cirolanid isopod from Madison Cave, Virginia. Internat. J. Speleology, 1: 229-236, 8 pls.
Burkhardt, C.
1906. Geologic de la Sierra de Mazapil et Santa Rosa (México). 10th Internat. Geol. Cong. México, Guidebook 26: 1-40, 2 maps.
1930. Etude synthetiqtie sur le Mesozoique Mexicain. Sec. Paleont. Suisse, 40: 1-280 .
Cabrera, A. L.
1955. Latin America. In, Arid Zones Research VI, Plant Ecology, Reviews of Research. U.N.E.S.C.O. PP. 77-113.
Carozzi, A. V.
1962. Observations on algal biostromes in the Great Salt Lake, Utah J. Geol., 70: 246-252.
Cole, G. A.
1963. The Ai-nerican Southwest and Middle America. In, Limnology in North America. Univ. Wisc. Press. PP. 393-434.
1968. Desert L;mnology. In, Desert Biology, Volume 1. Academic Press. Inc., New York. PP. 423-486.
Cole, G. A., and M. C. Whiteside
1965. Kiatutblana - A limnological appraisal. If. Chemical factors and biota. Plateau, 38: 36-48.
Cole, G.A., and W. L. Mincklev
1966. Speocirolana thermydronis, a new species of cirolanid isopod crustacean from central Coahuila, México. Tulane Stud. Zool., 13: 17-22.
1968. "Anomalous" thermal conditions in a hypersaline inland pond. Trans. Ariz. Acad. Sci., 5: IL05-107.
1969. Occurrence of the asellid subfamily Stenasellinae (Crustacea, Isopoda) in the Western Hemisphere, with description of a new genus and species. Internat. J. Speleol., in press.
Conant, R.
1963. Semi-aquatic snakes of the genus Thamnophis from the isolated drainage system of the Río Nazas and adjacent areas in México. Copeia, 1963:473-479.
Contreras Arias, A.
1942. Mapa de las provincias climatol6gicas de la Republica Mexicana. Secretaria de Agricult. y Fomento, Dir. de Geograf., Meteorol., y Hidrolog., Inst. Geogr., México, D. F.
Damon, P. E., C. V. Haynes, and G. A. Cole
1964. Carbon-14 content of aquatic plants, Montezuma Well, Arizona. Trans. Amer. Geophys. Union, 45: 117.
Deevey, E. S., Jr.
1957. Limnological studies in Middle America, with a chapter on Aztec limnology. Trans. Conn. Acad. Sci. Arts, 39:213-328, 4 Pls.
Drake, R. J.
1951. Humboltiana taylori, new species, from northern Coahuila. Rev. Soc. Malac. "Carlos de la Torre," 8: 93-95.
Dunn, J. R.
1953. The origin of the deposits of tufa in Mono Lake. J. Sediment. Petrol., 23:18-23.
Germand, K. W.
1939. Lake basins of the Llano Estacado. Compass of Sigma Gamma Epsilon, 20 : 162-165.
Gilmore, R. M.
1947. Report of mammal bones from archeological sites in Coahuila, México. J. Mamm., 28: 147-165, 1 Pl.
Coldman, E. A.
1951. Biological investigations in México. Smithsonian Misc. Coll., 115: i-xiii, 1-476, frontis., 70 Pls., 1 map.
Goldman, E. A., and R. T. Moore
1945. The biotic provinces of México. J. Mamm., 26 : 347-360.
Gregory, T.
1938. Lion in the Carmens. Chicago Notices, 1: 70-81, 110-120, 1 map.
Harrington, E. R.
1948. Craters and crater springs of the Río Salado. J. Geol., 56: 182-185, 2 Pls.
Herrick, C. L.
1904. Lake Otero, an ancient salt lake basin in southeastern New México. Amer. Ceol., 34: 174-189.
Hevly, R. H., and P. S. Martin
1961. Geochronology of Pluvial Lake Cochise, southern Arizona. I. Pollen analysis of shore deposits. J. Ariz. Acad. Sci., 2: 24-31.
Hubbs, C. L., and R. R. Miller
1948. The zoological evidence. Correlation between fish distribution and hydrographic history in the desert basins of Western United States. Bull. Univ. Utah, 38, Biol. Ser., 10: 17-166.
1965. Studies of cyprinodont fishes. XXII. Variation in Lucania parva, its estabIishment in Western United States, and description of a new species from an interior basin in Coahuila, México. Misc. Ptibi. Mus. Zool., Univ. Mich., 127: 1-111, 2 Pls .
Hubbs, C. L., C. S. Bien, and H. E. Suess
1965. La Jolla radiocarbon measurements. IV. Radiocarbon, 7: 66-117.
Hobbs, C.
1954. Corrected distributional records for Texas fresh-water fishes. Tex. J. Sci., 6: 277-291.
Hobbs, C., and W. H. Brown
1956. Dionda diaboli (Cyprinidae), a new minnow from Texas. SW Nat., 1: 69-77.
Hubbs, C., R. A. Kuebne, and J. C. Ball
1953. The fishes of the upper Guadalupe River, Texas. Tex. J. Sci., 5:216-244.
Humphrey, W. E.
1956. Tectonic framework of northeast México. Trans. Gulf Coast Assn. Geol. Sec., 6: 26-35.
Imlay, R. W.
1936. Evolution of the Coahuila Peninsula, México. IV. Geology of the western part of the Sierra de Parras. Bull. Geol. Soc. Amer., 47: 1091-1152, 10 Pls.
Jaeger, E. C.
1957. The North American Deserts. Stanford Univ. Press. Johnston, I. M.
1943. Plants of Coahuila, eastern Chihuahua, and adjoining Zacatecas and Durango. I-II. J. Arnold Arboretum, 24: 306-339.
1944. Ibid., III-V, 25: 47-83, 113-182, 431-453.
Jones, J. C.
1914. The tufa deposits of the Salton Sink. In, The Salton Sea, a study of the geology, the floristics, and the ecology of a desert basin. Carnegie Inst. Wash., Publ. 193. PP. 79-83.
Jones, T.S.
1938. Geology of Sierra de la Pena and paleontology of the Indura formations, Coahuila, México. Bull. Ceol. Sec. Amer., 49: 69-150, 13 Pls.
Judson, S.
1950. Depressions of the northern portion of the southern High Plains of New México. Ibid., 61: 253-274.
Kalhnan, K.
1964. Genetics of tissue transplantation in isolated platyfish populations. Copeia, 1964: 513-522.
Keith, A.
1928. Structural symmetry in North America. Bull. Geol. Sec. Amer., 39: 321-394.
Kellum, L. B.
1932. Reconnaissance studies in the Sierra de jimulco, México. Ibid., 43: 543.-564. 1936. Evolution of the Coahuila Peninsula, México. III. Geology of the mountains west of the Laguna District. Ibid., 47: 1039-1090, 14 Pls.
Kellum, L. B., R. W. Imlay, and W. G. Kane
1936. Evolution of the Coahuila Peninsula, México. 1. Relation of structure, stratigraphy and igneous activity to an early continental margin. Ibid., 47: 969-1008, 3 PlS.
Kelly, W. A.
1936. Evolution of the Coahuila Peninsula, México. II. Geology of the mountains bordering the valleys of Acatita and las Delicias. Ibid., 47: 1009-1038, 13 Pls.
King, P. B.
1935. Outline of structural development of Trans-Pecos Texas. Bull. Amer. Assoc. Petrol. Geol., 19: 221-261.
King, R. E.
1934. The Permian of southwestern Coahuila, México. Amer. J. Sci., Ser. 5, 27: 98- 112.
Kottlowski, F. E.
1958. Lake Otero - second phase in formation of New México's gypsum dunes. Bull. Geol. Sec. Amer., 69: 1733-1734 (abstr.).
Legler, J. M.
1960. A new subspecies of slider turtle (Pseudemys scripta) from Coahuila, México. Univ. Kans. Publ., Mus. Nat. Hist., 13: 73-84, 4 Pls.
Leopold, A. S.
1950. Vegetation zones of México. Ecology, 31: 507-518.
Maldonado-Koerdell, M.
1958. Nomenclature, bibliografia, y correlaci6n de las formaciones continentales (y algunas marinas) de Mesozoico de México. Bull. Assoc. Mex. Geol. Petrol., 10: 287-308.
1964. Geohistory and paleography of Middle America. In, Handbook of Middle American Indians, Volume 1. Natural Environments and Early Cultures. Univ. Tex. Press. PP. 3-32.
Martin, P. S.
1958a. Pleistocene ecology and biogeography of North America. In, Zoogeography. A.A.A.S. Publ. 51. PP. 375-420. 1958b. Biogeography of reptiles and amphibians in the Gomez Farias region, Tamaulipas, México. Misc. Publ. Mus. Zool., Univ. Mich., lol: 1-102, 7 Pls.
Martin, P. S., and B. E. Harrell
1957. The Pleistocene history of temperate biotas in México and eastern United States. Ecology, 38: 468-480.
Martin, P. S., and P. J. Mehringer, Jr.
1965. Pleistocene pollen analysis and biogeography of the southwest. In, The Quaternary of the United States. Princeton Univ. Press. PP. 433-451.
McBride, G. M.
1923. The land system of México. Amer. Geogr. Sec., Res. Ser., 12: i-Xii, 1-204.
Meek, S. E.
1904. The fresh-water fishes of México north of the Isthmus of Tehuantepec. Field Columbia Mus., Publ. 93 (Zool. Ser.), 5: i-lxiv, 1-252, 15 Pls.
Meinzer, 0. E., and F. C. Kelton
1913. Geology and water resources of the Sulphur Springs Valley, Arizona. U. S. Geol. Surv., Water Suppl. Pap., 320: 9-213.
Miller, R. R.
1948. The cyprinodont fishes of the Death Valley system of eastern California and Southwestern Nevada. Misc. Publ. Mus. Zool., Univ. Mich., 68: 1-155, 15 Pls, 1 map.
1958. Origin and affinities of the freshwater fish fauna of western North America. In, Zoogeography. A.A.A.S. Publ. 51. PP. 187-222 .
1961. Man and the changing fish fauna of the American Southwest. Pap. Mich. Acad. Sci., Arts, Lett., 46: 365-404.
1968. Two new fishes of the genus Cyprinodon from the Cuatro Ciénegas basin, Coahuila, México. Oce. Pap. Mus. Zool., Univ. Mich., 659: 1-15.
Miller, R. R. and W. L. Minckley
1963. Xiphophorus gordoni, a new species of platyfish from Coahuila, México. Copeia, 1963: 538-546.
Milstead, W. W.
1960. Relict species of the Chihuahuan Desert. SW Nat., 5: 75-88.
Minckley, W. L.
1962. Two new species of fishes of the genus Gambusia (Poeciliidae) from northeastern México. Copeia, 1962: 391-396.
Minckley, W. L., and M. Itzkowitz
1967. Ecology and effects of intestinal gas accumulation in a pupflsh (genus Cyprinodon). Trans. Amer. Fish. Sec., 96: 216-218.
Minckley, W. L., and G. A. Cole
1968a. Preliminary limnologic information on waters of the Cuatro Ciénegas basin, Coahuila, México. SW Nat., 13: 421-431.
1968b. Speocirolana thermydronis from northern México, re-discovery, habitat,
and supplemental description. Tulane Stud. Zool., 15: 2-4.
Minckley, W. L., and J. E. Deacon
1968. Southwestern Fishes and and the enigma of "endangered species." Science, 159: 1424-1432.
Mitchell, R. W.
1956. Winter invertebrate metazoa of Goose Lake, Muleshoe Wildlife Reserve, Texas. SW Nat., 1: 6-15.
Moore, G. A.
1968. Fishes. In, Vertebrates of the United States. McGraw-Hill Book Co. Pp.21-165.
Morrison, R. B.
1965. Quaternary geology of the Great Basin. In, The Quaternary of the United States. Princeton Univ. Press. Pp. 265-286.
Müller, C. H.
1947. Vegetation and climate of Coahuila, México. Madroiio, 9: 33-57.
Mulleried, F. K. G.
1942. The Mesozoic of México and northwestern Central America. Proc. 8th Amer. Sci. Congr., 4: 124-147.
Murray, G. E.
1961. Geology of the Atlantic and Gulf Coastal Province of North America. Harper and Brothers, New York.
Olive, W. W.
1955. Subsidence troughs in the Castile Anhydrite of the gypsum plain, New México and Texas. Bull. Geol. Sec. Amer., 66 (Pt. 2): 1604 (abstr.).
Parker, J. M., and C. J. Whitfield
1941. Ecological relationships of playa lakes in the southern Great Plains. J. Amer. Sec. Agron., 33: 125-129.
Pennak, R. W.
1958. Some problems of freshwater invertebrate distribution in the western states. In, Zoogeography. A.A.A.S. Publ. 51. PP. 223-230.
Pearse, A. S. (Editor)
1936. The Cenotes of YucatAn, a zoological and bydrographic survey. Carnegie Inst. Wash., Publ. 457: i-iii, 1-304, 8 Pls.
Porter, W. W.
1932. The Coahuila Piedmont, a physiographic province in northeastern México. J. Geol., 40: 338-352.
Reed, E. L.
1930. Vegetation of the playa lakes in the Staked Plains of western Texas. Ecology, 13: 597-6oo.
Robles Ramos, R.
1942. Orogenisis de la RepAblica Mexicana en relaci6n a so relieve actual. Irrigación en México, 23: 6-61.
Rodriguez Gonzáles, J.
1926. Geografia del Estado de Coahuila. Sec. Ediciòn y Libreria Franco-Americana, México, D. F.
Rosen, D. E., and R. M. Bailey
1963. The poecillid fishes (Cyprinodontiformes), their structure, zoogeography, and systematics. Bull. Amer. Mus. Nat. Hist., IL26: 1-176, 2 Pls.
Schmidt, K. P., and D. W. Owens
1944. Amphibians and reptiles of northern Coahuila, México. Pub]. Field Mus.
Nat. Hist. (ZOOI.) 29: 97-115.
Scbuchert, C.
1935. Historical Geology of the Antillean-Caribbean Region, or the Lands Bordering the Gulf of México and the Caribbean Sea. John Wiley and Sons, New York.
Shields, L. M.
1-956. Zonation of vegetation within the Tularosa basin, New México. SW Nat., 1: 49-68.
Shreve, F.
1942. Grassland and related vegetation in northern México. Madrofio, 6: Igo-198. 1944. Rainfall of northern México. Ecology, 25: 105-111.
Singewald, Q. D.
1936. Evolution of the Coahuila Peninsula, México. V. Igneous phenomena and geologic structure near Mapimi. Bull. Geol. Sec. Amer., 47: 1153-l176, 5 Pls.
Smalley, A. E.
1964. A new Palaemonetes from México. Crustaceana, 6: 229-232.
Taylor, D. W.
1966. A remarkable snail fauna from Coahuila, México. Veliger, 9: 152-228,
1 3. Pls.
Taylor, D. W., and W. L. Minckley
1966. New world for biologists. Pacific Disc., 19: 18-22.
Taylor, W. K.
1966. Avian records from central Coahuila, M6xico, primarily from the Cuatro Ciénegas area. SW Nat., 11: 136-137.
Taylor, W. P., W. B. McDougall, C. C. Presnall, and K. P. Schmidt
1946. The Sierra del Carmen in northern Coahuila. Tex. Geogr. Mag., 1946: 11-22, i map.
Taylor, W. W.
1956. Some implications of the Carbon-14 dates from a cave in Coahuila, México. Bull. Tex. Archeol. Sec., 27: 215-234, 3 Pls.
1962. Tethered nomadism and water territoriality: an hypothesis. Actas y Memorias, 35th Congr. Internat. de Americanistas, 1962: 197-203.
1966. Archaic cultures adjacent to the northeastern frontiers of Mesoamerica. In, Handbook of Middle American Indians, Volume 4, Archaeological Frontiers and External Connections. Univ. Tex. Press. PP. 59-94.
Thayer, W. N.
1916. The physiography of México. J. GeOl., 24: 61-94.
Urban, E. K.
1959. Birds from Coahuila, México. Univ. Kans. Pub]., Mus. Nat. Hist., 11: 443-516.
Vivó Escoto, J. A.
1964. Weather and climate of México and Central America. In, Handbook of Middle American Indians, Volume i, Natural Environments and Early Cultures. Univ. Tex. Press. Pp. 187-215.
Webb, R. G., and J. M. Legler
1960. A new soft-shelled turtle (genus Trionyx) from Coahuila, México. Univ. Kans. Sci. Bull., 40: 21-30, 2 Pls.
Webb, R. G., W. L. Minckley, and J. E. Craddock
1963. Remarks on the CoahuiIan box turtle, Terrapene coahuila (Testudines, Emydidae). SW Nat., 8: 89-99.
Weidie, A. E., and G. E. Murray
1967. Geology of Parras basin and adjacent areas of northeastern México. Amer.
Assn. Petrol. Geol. Bull., 51: 678-695.
Welch, P. S.
1948. Limnological Methods. Blakiston Co., Philadelphia.
West, R. G.
1964. Surface configuration and associated geology of Middle America. In, Handbook of Middle American Indians, Volume 1, Natural Environments and Early Cultures. Univ. Tex. Press. PP. 33-83.
Wilson, M. S., and H. C. Yeatman
1959. Harpacticoida. In, Fresb-water Biology, 2nd Edition. John Wiley and Sons, New York. Pp. 815-868.
Williams, S. C.
1968. Scorpions from northern México: five new species of Vejovis from Coahuila, México. Occ. Pap. Calif. Acad. Sci., 68: 1-24.