THE HYDROBIID SNAILS (GASTROPODA: RISSOACEA) OF THE CUATRO CIÉNEGAS BASIN: SYSTEMATIC RELATIONSHIPS AND ECOLOGY OF A UNIQUE FAUNA
ROBERT HERSHLER
Edwards Aquifer Research and Data Center
Southwest Texas State University
San Marcos, Texas 78666-4615
ABSTRACT
Results of the study of the morphology, systematics, and ecology of the Cuatro Ciénegas hydrobiids are given. Contrary to previous thought, no subfamilies of hydrobiids are endemic to the basin: all taxa studied belong to either the Nymphophilinae or Littoridininae, subfamilies widespread throughout North America. Of the nine genera (five endemic) and 13 species (nine endemic) found while sampling a large portion of the basin drainage, one genus and three species are new (but will be described elsewhere), and two species are new to the basin. The six nominal species of Mexipyrgus are reduced to one, M. churinceanus.
The diverse fauna is partitioned among three habitat types: species with large, thickened shells inhabit large springs and their outflows; minute, blind, unpigmented species are restricted to smaller groundwater outlets; and a third set of species inhabits smaller streams of the basin. Within large springs, micro-habitat partitioning occurs as separate species predominate either in soft sediment, on aquatic vegetation, or on travertine. At least one species reproduces year-round in the thermal waters of large springs.
Phenetic and phyletic analyses show that three of five endemic genera closely resemble non-endemics found in the basin. The close similarity of endemics to non-endemics, lower level of endemism than once thought, lack of marked differentiation within the basin, and close proximity of the basin drainage to outside waters suggest a local and recent origin for the endemic taxa.
RESUMEN
Se reportan resultados del estudio de la morfología, sistemática, y ecología de los hydrobioides de Cuatro Ciénegas. Contra lo que se habia pensado, ninquna subfamilia de hydrobioids es endémica a la cuenca: todos los taxa investigados pertenecen a Nymphophilinae ó Littoridininae, subfamilies extendidas por todo Norte America. De los 9 géneros (5 endémicos) y 13 especies (9 endémicas) encontrados en muestras tomadas sobre una porción grande de la cuenca, 1 género y 3 especies son nuevas, y 2 especies son nuevas para la cuenca. Las 6 especies nominates de Mexipyrgus estan reducidas a 1, M. churinceanus.
La fauna diversa está repartido entre 3 tipos de hábitats: las especies con conchas grandes y gruesas habitan manantiales grandes y sus desagues; las especies diminutas, ciegas, y sin pigmentación estan restringidas a salidas pequeñas de aguas subterráneas; y el tercer grupo de especies habita arroyos pequeños. Dentro de los manantiales grandes se ve la repartición de microhábitat donde especies distintas predominan en sedimento suelto, en vegetación acuática ó en travertino. Por lo menos una especie se reproduce durante todo el año en las aguas termales de los manantiales grandes.
El análisis fenotípico y filético muestra que 3 de los 5 géneros endémicos son muy parecidos a los no endémicos encontrados en la cuenca. La semejanza casi completa de los endémicos a los no endémicos, el nivel mas bajo de endemismo que se habia pensado, la falta de diferenciación dentro de la cuenca y la proximidad cercana del drenaje de la cuenca a aguas afuera indican un origen local y reciente de los taxa endémicos.
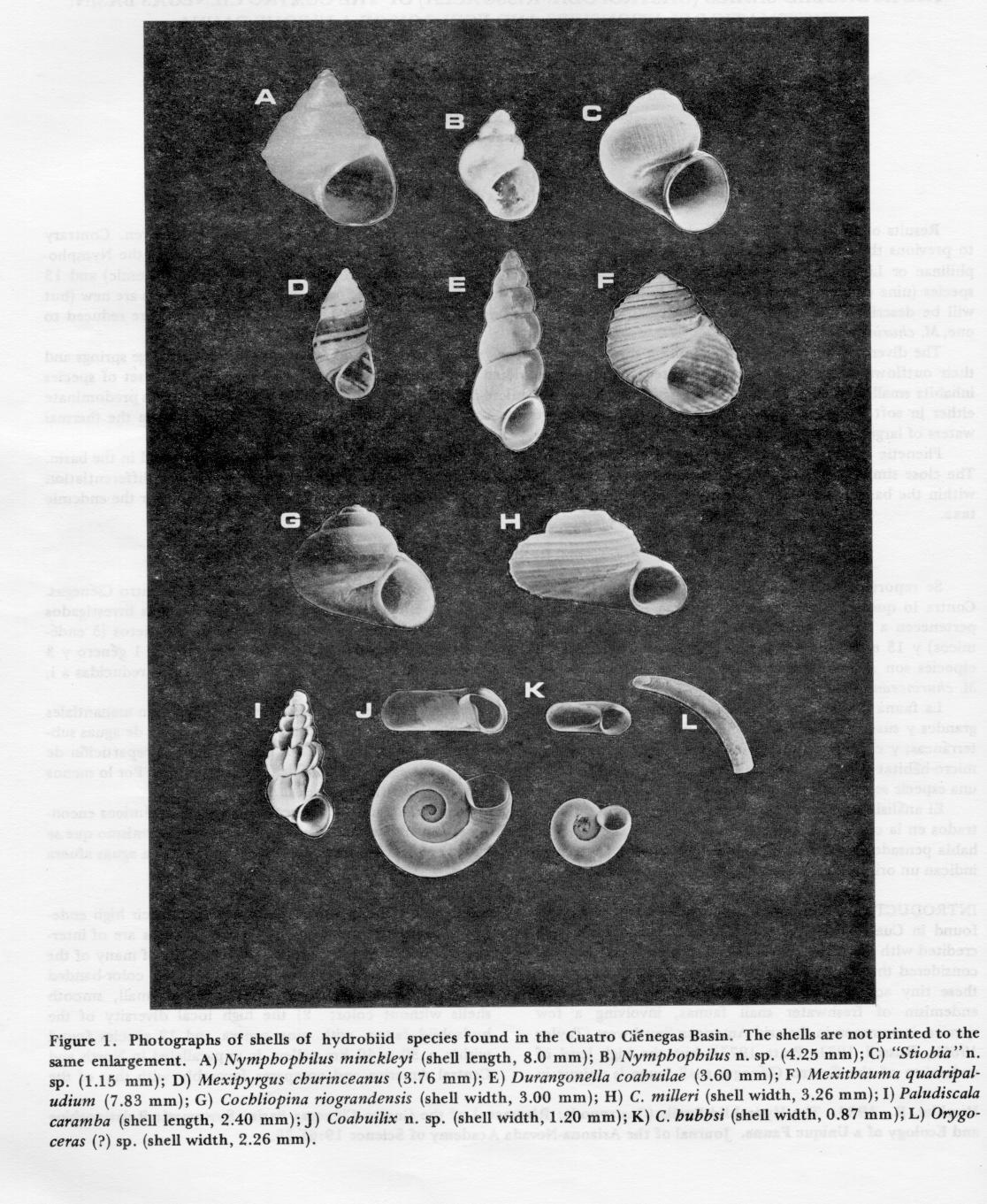
INTRODUCTION.-Of the various groups of organisms found in Cuatro Ciénegas, the hydrobiid snails have been credited with the highest level of endemism. Taylor (1966a) considered three subfamilies, five genera, and 12 species of these tiny aquatic snails as endemic to the basin. Local endemism of freshwater snail faunas, involving a few species, is common in the arid American Southwest (Taylor 1966b, Russell 1971, Pratt 1977), but the high level of endemism of the Cuatro Ciénegas snail fauna is unique in North and Central America. Apart from their high endemism, the hydrobiid snails of Cuatro Ciénegas are of interest for the following reasons: 1) the shells of many of the species are relatively large, sculptured, or color-banded (Fig. 1) , whereas most hydrobiids have small, smooth shells without color; 2) the high local diversity of the hydrobiid fauna, with nine genera and 12 species found within a 30 x 40 km valley, is unparalleled in North and Central America and compares favorably with that of the large hydrobiid faunas of ancient lakes (Table 1); and 3) differentiated populations of snails distributed among the insular springs of the valley offer an opportunity to study evolution in a "natural laboratory" (Taylor and Minckley 1966).
{kind=link}
Early classifications of hydrobioids (i.e., those rissoacean snails that resemble Hydrobia in morphology of either shell, penis, operculum, or radula) (Berry 1943; Morrison 1949), including that originally offered for the Cuatro Ciénegas fauna (Taylor 1966a), were based on these characters and a few other external morphological features (see Table 2). It now is known that such characters have converged in rissoacean taxa that are not closely related on the basis of overall soft-part anatomy (Davis 1979). Such convergences are rampant among hydrobiid snails and study of entire soft-part anatomy is necessary both to recognize these convergences and to clarify systematic relationships (Davis 1979). Endemic freshwater snail faunas frequently include taxa with thickened, sculptured, or otherwise-peculiar shells (Annandale 1919, Vermeij and Covich 1978) that represent adaptations to local ecological conditions.
Classifications weighted in favor of such shell features may inflate the uniqueness of such taxa. Some of the endemic snails of Lake Tanganyika, for instance, have thickened, sculptured, "marine-like" shells and once were considered to be oceanic relicts and classified as members of modern or Jurassic marine families (see Pilsbry and Bequaert 1927; Yonge 1938). Later anatomical studies showed that they were modified members of widespread freshwater cerithiacean families (Leloup 1953): the peculiar shells masked a more conventional soft-part anatomy.
The original description of the Cuatro Ciénegas hydrobiids by Taylor (1966a) stands as a fine contribution for that time period and stimulated my own research. Yet in light of the above, the classification offered for the snails is suspect because it was based largely on shell and external features. Additionally, within the data set used, the numerous subfamilies described do not have distinctive sets of features (see Thompson 1968; Davis 1979). The descriptions of species are suspect also because they were based on samples from only twelve localities (Taylor 1966a): six allopatric species of Mexipyrgus were described from a total of seven localities.
A detailed presentation of the morphology of the Cuatro Ciénegas hydrobiids, together with a systematic revision of the fauna (including descriptions of new taxa), is presented elsewhere (Hershler 1983). The purposes of this paper are: 1) to summarize changes in the classification of the snails and to discuss new taxa (and those new to the basin); 2) to present sampling data and to discuss patterns of distribution of the hydrobiid species among the aquatic habitats of the basin and other aspects of their ecology; and 3) to discuss the origin, evolution, and endemism of the hydrobiid fauna.
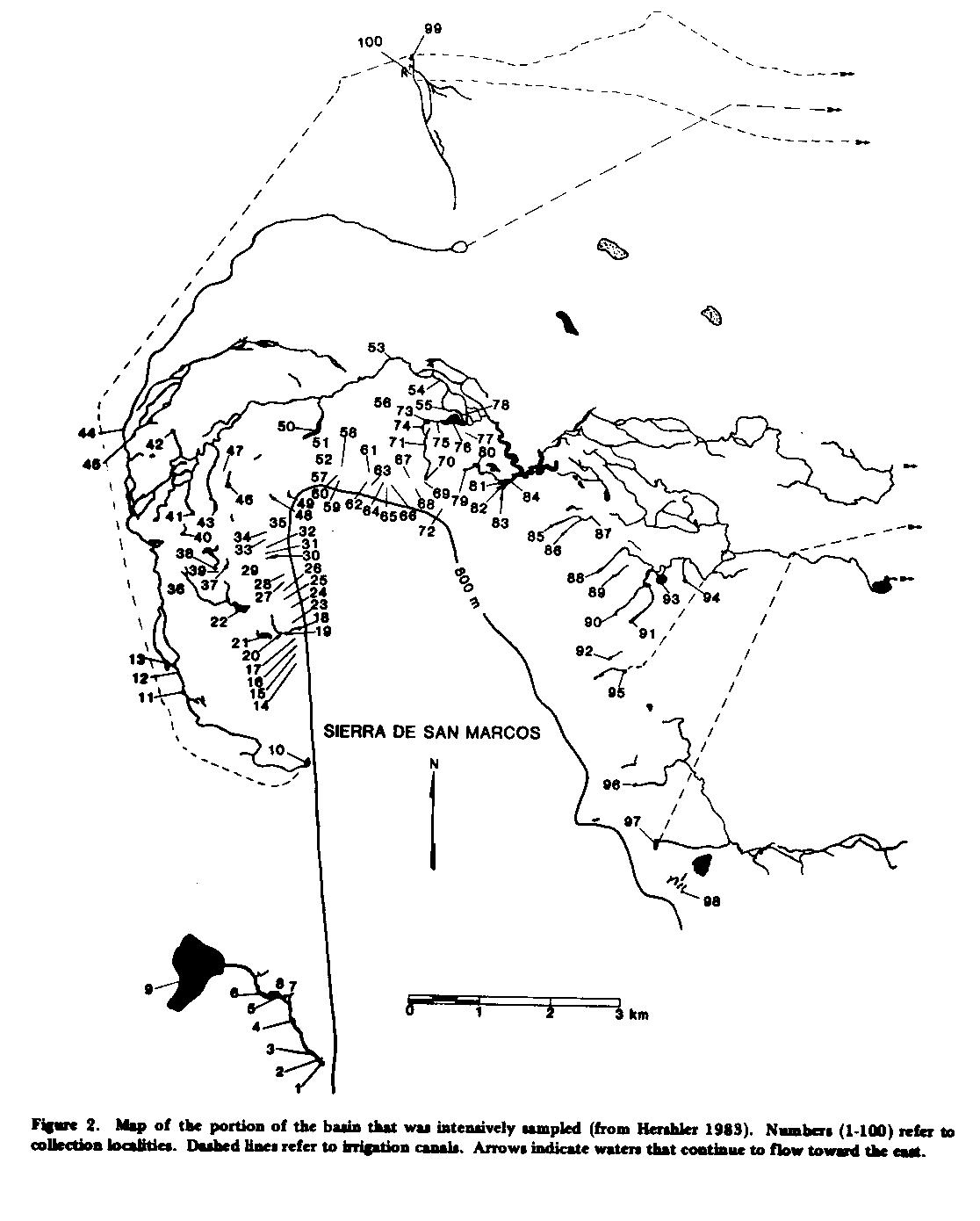
METHODS.-Snails were collected from 103 localities in the basin. One hundred localities were concentrated in an arc surrounding the northern tip of the Sierra de San Marcos (Fig. 2) and encompassed parts of four of the five-seven drainages of the basin (Minckley 1969; LaBounty 1974). Soft sediments, largely consisting of snail copropel, were sampled using a fine hand sieve. Bunches of vegetation (Clara, Utricularia, Nymphaea) were collected at random, washed, and the dislodged snails then picked from the residue under a dissecting microscope. Snails were picked from individual travertine pieces and blocks using fine forceps. To collect the tiny snails that frequent groundwater outlets, ordinary domestic (cotton) mops were placed into small springheads to serve as colonization substrates (method suggested by W. L. Minckley). Mops were periodically removed, washed, and the snails picked from the residue under a dissecting microscope. Dissection methods and other techniques of morphologic study are described elsewhere (Hershler 1983).
{kind=link}
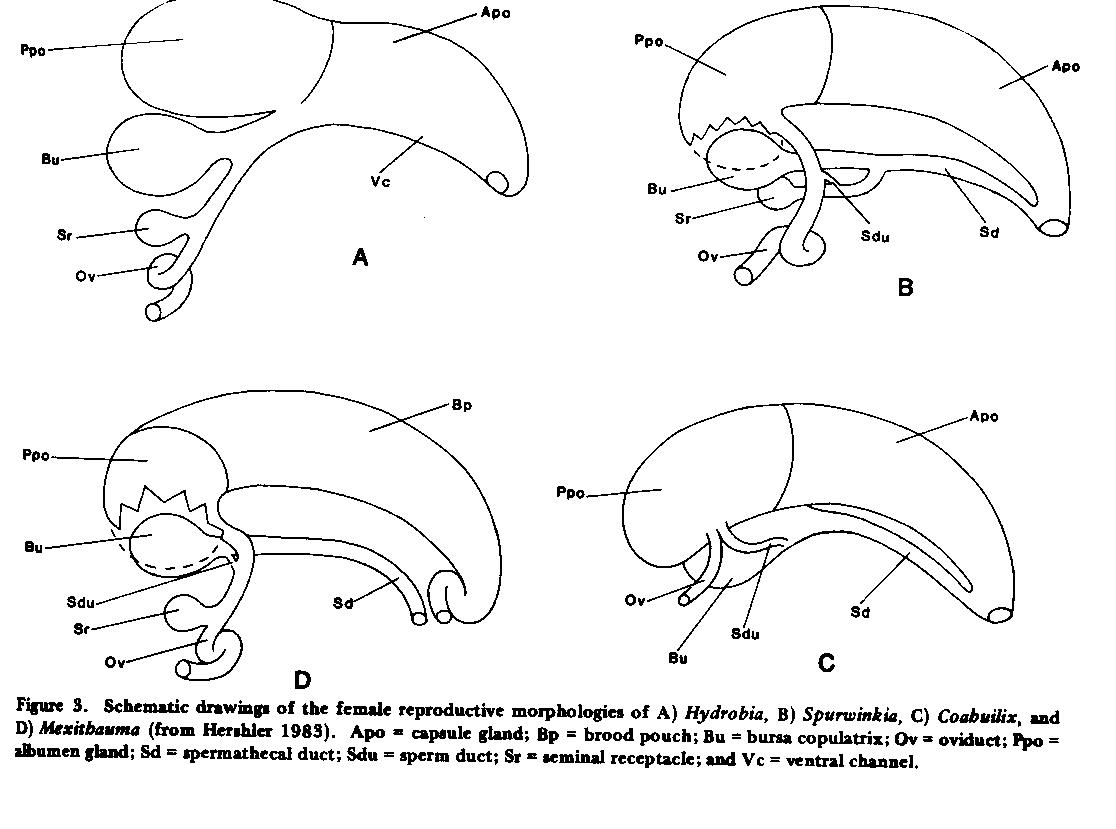
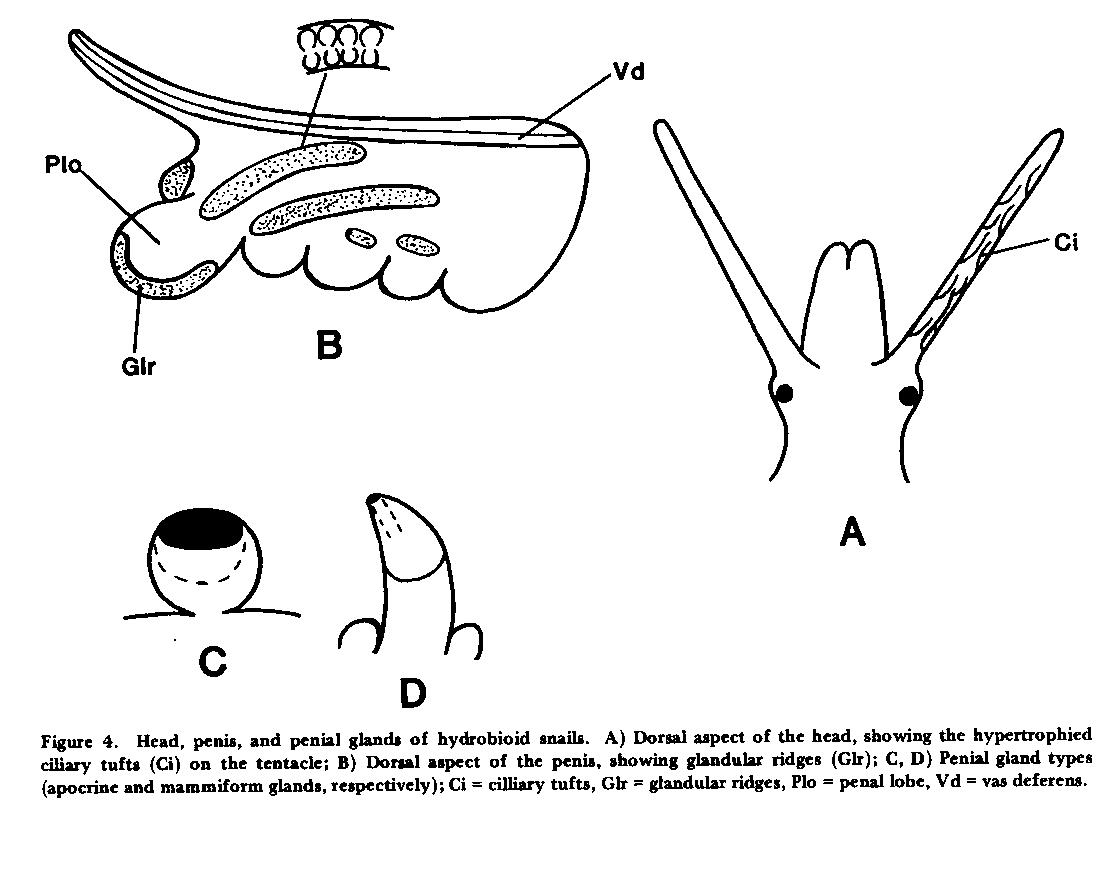
RESULTS.-Changes in classification. Recent anatomical studies have demonstrated that hydrobioid snails represent a polyphyletic assemblage that includes taxa belonging to the Hydrobiidae and Pomatiopsidae, families characterized by divergent anatomical groundplans (Davis 1979). Among the Hydrobiidae, two basic anatomical groups are seen (Davis et al. 1982, Hershler 1983). One group includes the Hydrobiinae and Nymphophilinae (sensu Thompson 1979, Davis et al. 1982), in which sperm are received into the pallial oviduct of the female (Apo + Ppo, Fig. 3a ) and travel along an internal ciliated channel (Vc) of the capsule gland (Apo) towards the sperm-receiving bursa copulatrix (Bu) and seminal receptacle (Sr). Nymphophilines are characterized by a penis with glandular ridges (Glr, Fig. 4B ), elevated regions with paired rows of small glands discharging through a central slit; whereas species in the Hydrobiinae have a penis lacking specialized glands. In the other group, the Littoridininae (sensu Davis et al. 1982), sperm enter a spermathecal duct (Sd, Fig. 3B ) of the female, which is at least partly separate from the pallial oviduct (wholly separate in some taxa, Fig. 3D ), and travel along this duct: the pallial oviduct has no ciliated sperm groove in the ventral channel. This penis of littoridinines lacks glandular ridges, but may have other specialized gland types ( Figs. 4C , D ). Littoridinines also differ from the Nymphophilinae-Hydrobiinae in that their cephalic tentacles often have complex, hypertrophied, surficial ciliary tracts (Ci, Fig. 4A ) , while the tentacles of the latter lack these complex tracts. This subfamilial classification of the Hydrobiidae, while admittedly based on few characters, does place emphasis on a major difference in overall anatomical groundplan (i.e., ventral channel versus spermathecal duct). This classification is deemed tentative, not only because at least 90% of the North and Central American hydrobiid species are unknown in terms of anatomy, but also because recent work (Ponder 1983) suggests that character-states of the female reproductive system (such as the above) may also have arisen iteratively in the evolution of various rissoacean groups.
{kind=link}
{kind=link}
Thompson (1979) demonstrated that Nymphophilus, endemic to Cuatro Ciénegas and previously considered the sole genus in this subfamily (Taylor 1966a), was similar in anatomy to hydrobiid snails from other parts of North America and added several genera to this subfamily. My studies indicate that all Cuatro Ciénegas hydrobiids studied can be placed in the Nymphophilinae or the Littoridininae (also widespread with numerous taxa): there are no subfamilies endemic to the basin. My classification of the Cuatro Ciénegas hydrobiids is contrasted with the original classification in Table 3. The purported distinctive features of the three nominal subfamilies erected by Taylor (1966a), which were considered endemic to the basin (Table 1), are unconvincing. For example, the penis of Nymphophilus has glandular ridges as do those of all nymphophilines, and all hydrobiids have the vas deferens in a ridge (of varying height) on the floor of the mantle cavity (Thompson 1968, 1979). The reflected pallial oviduct in Mexithauma is found in many hydrobiids and is associated with a viviparous reproductive habit (Hershler 1983). Pigment streaks on tentacles also are common among hydrobiid snails (Fretter and Graham 1962, Hershler and Davis 1980). Lamelliform costae on the shell, seen in Paludiscala, also are found in unrelated taxa (e.g., Lanzaia).
The six nominal species of Mexipyrgus are reduced to one, M. churinceanus Taylor ( Fig. 1D ). The nominal species were described on the basis of minor differences in shell features. I gathered morphological data on specimens from 33 populations and found that while patterns of geographical variation were evident, the purported differences in shell features among nominal species often were blurred in given populations, or even intergrade, and distinct species thus could not be recognized (Hershler 1983).
New taxa. A list of the hydrobiid species found in the basin is given in Table 4. The new taxa will be described elsewhere (Hershler 1983). Coahuilix n. sp. (Fig. 1J) is much larger than C. hubbsi, and differs in other shell features as well when sympatric. Nymphophilus n. sp. ( Fig. 1B ) is much smaller in size than N. minckleyi and lacks the pronounced basal keel of the shell seen in the latter. These two species are allopatric to one another: Nymphophilus n. sp. is known only from the southeast lobe of the basin, whereas N. minckleyi is restricted to the northeast and western lobes. "Stiobia" n. sp. ( Fig. 1C ) represents a new genus of Nymphophilinae (also found elsewhere in Mexico), similar to Stiobia (Thompson and McCaleb 1978).
Two species new to the basin were discovered during my collecting. Cochliopina riograndensis ( Fig. 1G ), previously known from the nearby Río Salado de Nadadores and other Río Grande tributaries (Taylor 1966a), was collected in Laguna Santa Tecla in the southeastern lobe of the basin. Orygoceras(?) sp. ( Fig. 1 ), previously known from a single spring in western Texas (Taylor 1974), was discovered living in a single small spring in the basin. A fresh shell of this species has been found since at a second locality in western Texas (Davis 1983).
ECOLOGY.-As mentioned above, the hydrobiid fauna of Cuatro Ciénegas also is noteworthy for its high diversity. How are the 12 species distributed among the aquatic habitats of the basin? Most aquatic habitats in the basin are spring-fed, and vary greatly in basic type (i.e., spring pool, stream, marsh, lake), size, and temperature (Minckley 1969). For the purposes of describing the distribution of the various snail species, it is convenient to divide the springs into two classes: small springs, with spring pool areas of < 9 m2, whose outflows run as shallow streams for only tens of meters before terminating in a marsh or disappearing underground; and larger springs (Minckley 1969, Figs. 4 , 5 ), with spring pool areas in excess of 900 m2, whose outflows are large streams. Discussion is based on intensive collecting at Localities 1-100 ( Fig. 2 ): species not found in this area (Cochliopina riograndensis and Nymphophilus n. sp.) are discussed only briefly.
{kind=link}
Large springs and their outflows. The large springs have considerable micro-habitat diversity, including aquatic vegetation of several types (sedges, Nymphaea, Chara, Utricularia); soft sediment composed of snail copropel or an algal-detritus mixture, often mixed with small travertine pieces and shell debris; and large-sized travertine pieces ( Fig. 5 ). Mexipyrgus churinceanus, Mexithauma quadripaludium, and Nymphophilus minckleyi are commonly found in the large springs; while Durangonella coahuilae and Cochliopina milleri are much less common. This may be explained partly by the presence of molluscivorous cichlid fishes in the large springs (Deacon and Minckley 1974, Fig. 7 ): the former three snails have large and thickened shells, possibly affording resistance to the snail-crushing fish, whereas the latter two species are fragile-shelled.
{kind=link}
In all springs sampled, M. churinceanus was restricted to soft sediments (in which it burrows), sometimes achieving extremely high densities ranging to 40,000/m2 (data from box core sampling). Nymphophilus minckleyi and Mexithauma quadripaludium usually are found on hard substrates (plant or travertine). The massive foot and trochoid shell of these snails are presumably adaptations to a clinging mode of life. Nymphophilus minckleyi may also require a hard substrate on which to deposit its egg capsules (M. quadripaludium is viviparous). When N. minckleyi and M. quadripaludium coexist in a spring that has aquatic vegetation and travertine in abundance, each predominates on the substrate to which it is best adapted (the former on vegetation, the latter on travertine). Data on substrate utilization by the three species in Laguna Mojarral East, a large limnocrene (pool area of about 9,000 m2), are given in Table 5. Only M. churinceanus was common in soft sediments; N. minckleyi was common and M. quadripaludium rare on aquatic vegetation; and M. quadripaludium was three times as common as N. minckleyi on under surfaces of travertine blocks (the three substrates are shown in Fig. 5 ). Thus, in a large spring with micro-habitat diversity, each of the three species predominates in a separate micro-habitat. In springs with reduced micro-habitat diversity, either N. minckleyi or M. quadripaludium is rare or absent, and the species present often "switches" to the micro-habitat usually occupied by the other, suggesting at least a partly competitive basis for patterns of substrate use. In Laguna Churince, for instance, N. minckleyi was extremely rare, travertine was largely absent, and M. quadripaludium was extremely abundant on dense growths of Chara in the deeper portions of the spring.
The other two species occasionally found in large springs, Durangonella coahuilae and Cochliopina milleri, usually are restricted to soft sediment and aquatic vegetation, respectively. Cochliopina riograndensis and Nymphophilus n.sp. are on aquatic vegetation in large springs in the Southeastern lobe of the basin.
Prior to man-made alterations, most of the large springs gave rise to large streams that terminated in shallow playa lakes (Minckley 1969). In general, abundances of all of the above species, except Durangonella coahuilae, decrease downstream from a spring source. Several environmental changes occur downstream that may account for these decreases. Aquatic vegetation may disappear or become patchy and soft sediments frequently become covered with an algal layer. Water temperature fluctuations may become marked. At Locality 30 ( Fig. 2 ), for instance, water temperatures at the springhead varied from 31.0-34.5 degrees C, over a four-day period during the summer (maxi-mini thermometer readings), while 77 m downstream where water disappeared into a swallet, the temperatures varied from 20.6-37.8 degrees C during the same period. Salts become more concentrated downstream as well (Minckley and Cole 1968). In the Churince system (Localities 1-9, Fig. 2 ), M. churinceanus, M. quadripaludium, and N. minckleyi have not been found below the large pools (Locality 6), which differ greatly from the springhead in terms of substrate, water chemistry (Minckley and Cole 1968), and water temperature variations. Durangonella coahuilae, on the other hand, can tolerate these environmental changes and is abundant not only in these pools, but also in the playa lake, Laguna Grande (Locality 9).
Small springs and their outflows. Small springs also generally have warm water (29-33 degrees C). Riparian vegetation typically covers the springheads and sometimes part of the streams as well. Micro-habitat diversity is reduced relative to the large springs. Chara mats and soft sediments composed of mixtures of silt, organic mud and travertine are the only abundant substrates. Of the five species found in large springs, only D. coahuilae also is common in the smaller springs. One group of blind, unpigmented species (Paludiscala caramba, Coahuilix hubbsi, Coahilix n. sp., and Orygoceras[?] sp.) is restricted largely to heads of these small springs (and possibly subterranean waters), while a second group of species (D. coahuilae, "Stiobia" n. sp.) abounds not only in springheads, but also in the streams. Species of the second group have dark body pigment that may serve as "sunscreen" (shells are transparent, as are those of the first group), protecting the vital organs of the snails from the intense insolation of open stream reaches.
Data from the abundances of snail species in mop samples from 38 springheads are given in Table 6. At a given springhead, several mop samples were taken at varying intervals during 1980 and 1981. Snails were picked from each sample until a total of 100 individuals was found. Relative abundances varied little among samples of a given springhead, and data were pooled for each spring. Paludiscala caramba was by far the most common species in springheads (Table 7). Of the other blind species, both Coahuilix hubbsi and Coahuilix n. sp. were less common, and only a few specimens of Orygoceras (?) sp. were found in unsorted samples from a single spring. Proof is lacking that the blind, unpigmented species inhabit subterranean waters as well; the mops may be colonized not only by snails washed from underground, but also by surface-dwelling snails living in the springhead (such as Durangonella coahuilae and "Stiobia" n. sp.). To distinguish between these two sets of snails, a fine mesh net could be capped over the groundwater outlet, filtering snails from the watering stream, but not allowing surface-dwellers to crawl in. This method has been successfully used to sample faunas of artesian wells (Holsinger and Longley 1980).
While blind species were never found in the open reaches of stream, they occasionally extended below springheads when channels were covered by riparian vegetation (mimicking a subterranean environment). At Locality 63, a small thermal (33-35 degrees C) spring issued into a pool (Brown 1974, Fig. 5 ), then flowed 170 m as a shallow stream before entering a marsh. Paludiscala caramba was abundant on plant and travertine surfaces for the upper 83 m of stream, which had complete vegetative cover. Below 83 m, vegetative covere ended and no P. caramba were found despite intensive collecting that yielded quantities of Durangonella coahuilae and "Stiobia" n. sp. At Locality 67, nine mop samples taken 3 m below the springhead, where riparian vegetation completely covered the stream, yielded 67 P. caramba, 520 C. hubbsi, 10 Coahuilix n. sp., as well as quantities of the pigmented species.
The two pigmented species, D. coahuilae and "Stiobia" n. sp., are abundant in both soft sediments and Chara mats of shallow streams. No clear-cut differences were seen in their micro-habitat usage.
Other aquatic habitats. Of all species of the basin, only Durangonella coahuilae is common in playa lakes, pools and marshes, habitats with sluggish water circulation and pronounced water temperature and chemistry fluctuations. For example, only D. coahuilae was collected from a series of groundwater-fed pools that fluctuated in size as the water table rose and fell (Localities 7, 8). During 1981 water varied from 9.5-35.6 degrees C in one of these pools; the snails disappeared only when the pool temporarily dried during drought. Durangonella coahuilae does not, however, extend into the saline waters of the basin.
In summary (Table 8), hydrobiid species of the basin are divided into three groups with differing habitat usages: those dwelling in large springs and their outflows, blind species restricted to groundwater outlets, and two species common in the outflows of small springs (one of which also occupies marshes and playa lakes). Predominant species in large springs usually occupy different micro-habitats. As the fauna is large, partitioning of habitats occurs as expected, with no more than three or four species common at any locality. The diversity of habitats occupied by hydrobiids may be due partly to absence of other groups of gill-breathing snails (i.e., viviparids, pleurocarids). Adaptations of the species to their respective habitats and micro-habitats merit more study.
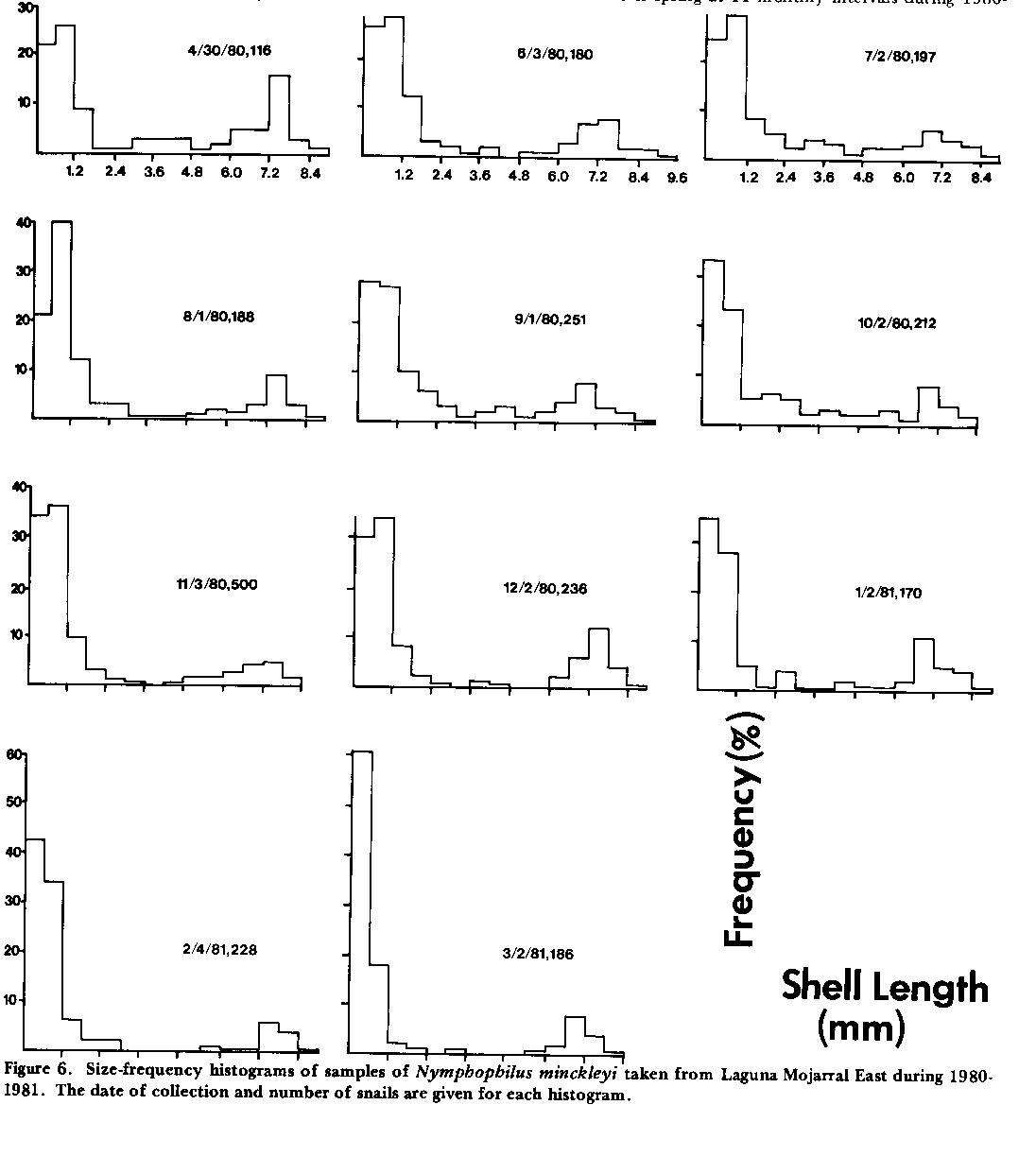
Reproductive ecology. Near-constancy of water temperatures in large spring pools of the basin is well documented (Minckley 1969, Arnold 1972) and is reflected in the apparent year-round reproduction of Nymphophilus minckleyi in Laguna Mojarral East ( Fig. 6 ). A large sample of Nymphaea (water lily) was collected from the south-west comer of this spring at 11 monthly intervals during 1980-1981 and washed to remove snails, which were then measured. Water temperatures throughout the year ranged from 26 to 32 degrees C. For all 11 samples, juveniles (shell length < 1.25 mm) comprised at least 48% of the total collection ( Fig. 6 ), with a March peak of about 80%. Individuals with mature gonads (seen in dissection), and egg capsules (on Nymphaea leaves) were common in all samples. Note the paucity of intermediate-sized (2.0-6.0 mm) snails in all samples ( Fig. 6 ), which may be due to intensive predation by cichlids in this spring. Tiny juveniles perhaps are overlooked by the fish whereas large adults may have thick enough shells to resist crushing forces of the molariform pharyngeal teeth of the cichlids. While comparable data are not available for other species of the basin, embryonic young were found in the brood sacs of all viviparous species (Durangonella coahuilae, Mexipyrgus churinceanus, Mexithauma quadripaludium, Cochliopina milleri) throughout the year.
{kind=link}
DISCUSSION.-Relationships among taxa. To confidently assess origins and relationships of a locally endemic fauna, a thorough knowledge is required of the systematics and distribution of that group over a much larger area. Such data are available, for example, for freshwater fishes of Mexico and adjacent regions, and thus the origins and affinities of various endemic fishes of Cuatro Ciénegas appear fairly clear (Miller 1978, Minckley 1978). Study of the hydrobioids of Mexico and adjacent regions lags far behind that of fishes; while numerous species have been described, neither anatomical information nor detailed distributional data are available for any species. Thus, while relationships among various members of the Cuatro Ciénegas fauna can be clarified, their relationships to members of other hydrobioid faunas remain unclear.
As mentioned above, the Cuatro Ciénegas hydrobiids (exclusive of Orygoceras [?] ) all belong to either the Nymphophilinae or Littoridininae (Table 3). Of the endemic genera, Nymphophilus belongs to the Nymphophilinae, whereas all other are Littoridininae. Nymphophilus and "Stiobia," the other nymphophiline genus in the basin, differ in at least 10 morphological features (Table 9), but many of those differences are probably correlates of their great size disparity (Nymphophilus = 3.5-8.4 mm shell length; "Stiobia" = 1.0-2.0 mm), and they may be closely related as they share several unusual features, notably the small number of glandular ridges on the penis.
A comparison was made among the six littoridinine genera of the basin involving 36 characters: six (17%) from the operculum and shell; seven (19%) from non-reproductive aspects of anatomy; eight (22%) from the male reproductive anatomy; and 15 (42%) from the female reproductive anatomy (see Hershler 1983 for a list of characters). Percentage difference among these taxa was determined by counting differences between pairs and dividing by total numbers of characters shared by the two taxa. A phenogram, based on simple averaging of differences between taxa, is given in Fig. 7A . Two pairs of very similar genera (< 29% difference) are each comprised of an endemic and non-endemic genus (Mexipyrgus, Durangonella; Mexithauma, Cochliopina). The other pair of genera (linking at 49% difference) are both endemic to the basin.
A hypothesis of phyletic relationships among Hydrobia (Hydrobiinae), the six Cuatro Ciénegas littoridinines, and Spurwinkia, the only other North American littoridinine known in terms of complete soft-part anatomy (Davis et al. 1982), is shown in Fig. 7B . Clades were defined on the basis of shared, unreversed, and presumably derived character states (numbers 1-10, Fig. 8 ) from aspects of soft-part anatomy (see Hershler 1983 for list). "A" represents a hypothetical, ancestral hydrobiid. The strong concordance between the grouping of Cuatro Ciénegas littoridinine genera in the phenogram and cladogram suggests that the groups constitute true clades. Distinguishing features of these three groups are given in Table 10. Note also that of the six genera, two (Coahuilix and Paludiscala) are egg-layers while the other four brood their young.
{kind=link}
Origin and endemism of the fauna. The idea that most of the endemic snails are ancient (tens of millions of years) relicts unrelated to other extant taxa of the region is prevalent in the literature (Taylor 1966a; Minckley 1969; Parodies 1969), but was based on classifications of the snails that inflated their seeming uniqueness. Unfortunately no fossil record, documenting timing of colonization(s) of and diversification within the basin, is known. However, my results suggest a more local and recent origin for the endemic snails.
I have shown that endemism is on a smaller scale than previously believed; no subfamilies are endemic to the basin. Of the five genera considered endemic to the basin, I suspect that two, Paludiscala and Coahuilix, are in fact not so restricted, but have not been recorded elsewhere because the techniques necessary to collect these minute snails from groundwater outlets generally are not used during fieldwork. Snails with shells resembling those of the above are known from the Edwards Aquifer of Texas (Pilsbry and Ferris 1906, Fullington 1978). Note that two of the three blind, unpigmented crustacean genera originally described from and considered endemic to groundwater outlets in the Cuatro Ciénegas Basin, Mexiweckelia and Mexistenasellus, were discovered later in cave waters in southern Mexico (Magniez 1972, Argano 1973, Holsinger 1973) and elsewhere (Cole, present symposium).
The basin drainage, while currently isolated, lies in close proximity to the Río Grande (= Río Bravo del Norte) drainage to the cast and has likely had repeated connections with these waters in the recent past (Minckley 1969, 1978). Taxa assignable to the Nymphophilinae and Littoridininae on the basis of penis type (i.e., Fontelicella and Pyrgophorus, respectively) are found in nearby waters of northern Mexico and southwest Texas.
The lack of marked differentiation of snail taxa within the basin may also suggest a young fauna; only three of the nine genera have a second species in the basin and in only one case (Coahuilix spp,) are congeners sympatric. It should be noted, however, that even over a long period of time speciation within the basin might be infrequent as the basin is small and its drainages have continually shifted through time (reducing the probability of continuous isolation of given springs).
The now-recognized smaller scale of endemism, similarity of endemics to non-endemics within the basin, proximity of the basin to outside drainage, and low level of intra-basin differentiation point to a recent (late Cenozoic?) and local origin for the endemic snail fauna. Note that the three genera that are definitely endemic to the basin (Nymphophilus, Mexipyrgus, and Mexithauma) are all inhabitants of large springs. Rapid in situ evolution of these genera (from the non-endemics they resemble) could have occurred as a result of intense cichlid predation within the large springs selecting for large, thickened shells, with soft parts changing less so. Rapid evolution of freshwater snail genera has been documented in several cases (Davis 1979, 1981; Stanley 1979).
ACKNOWLEDGMENTS.-My co-advisors, Drs. Steven M. Stanley and George M. Davis, provided constant encouragement during the tenure o f this project. Dr. Davis suggested the project to me, shared his vast knowledge of hydrobioid snails with me, and provided the space and facilities at the Academy of Natural Science of Philadelphia necessary for completion of the project. Drs. W. L. Minckley, G. Longley, I. Kornfield, and Mr. J. Landye discussed aspects of the project with me.
I thank the Mexican government for providing the necessary permits for collecting freshwater molluscs in their country. Fieldwork in Cuatro Ciénegas was aided by Dr. C. Dunn, Mr. D. Bereza and numerous local townspeople.
Financial support for the project came from Sigma Xi, the National Shell Club, the Theodore Roosevelt Memorial Fund, and the Geological Society of America. A Jessup Scholarship partly supported my stay at the Academy of Natural Science of Philadelphia.
Research for this paper was done in partial fulfillment for a Doctor of Philosophy degree at Johns Hopkins University, Baltimore, Maryland. I thank Drs. A. Metcalf, W. Ponder, and two anonymous reviewers for their comments on the manuscript.
LITERATURE CITED
ANNANDALE, N. 1919. The gastropod fauna of old lake beds in upper Burma. Rec. Geol. Soc. India 50: 209-240.
ARGANO, R. 1973. Mexistenasellus magniezi n. sp., a blind aquatic isopod from Veracruz, Mexico. Accademia Nazionale dei Lincei 171:97-103.
ARNOLD, E. A. 1972. Behavioral ecology of two pupfishes (Cyprinodontidae, genus Cyprinodon) from Northern Mexico. Unpubl. Ph.D. Dissert., Ariz. State Univ., Tempe.
BERRY, E. G. 1943. The Amnicolidae of Michigan: distribution, ecology and taxonomy. Misc. Publ. of the Mus. Zool., Univ. Mich. 57:1-68.
BOSS, K. J. 1978. On the evolution of gastropods in ancient lakes. Pp. 385-428. In: Pulmonates, Vol. 2A, Systematics, Evolution and Ecology (V. Fretter and J. Peake, Eds.), Academic Press, NY.
BROWN, W. S. 1974. Ecology of the aquatic box turtle Terrapene coahuila (Chelonia, Emydidae) in northern Mexico. Bull. Flor. State Mus., Biol. Sci. 19:1-67.
DAVIS, G. M. 1979. The origin and evolution of the gastropod family Pomatiopsidae, with emphasis on the Mekong River Triculinae. Acad. Nat. Sci. Phila., Monogr. 20:1-120.
DAVIS, G. M. 1981. Different modes of evolution and adaptive radiation in the Pomatiopsidae (Gastropoda: Mesogastropoda). Malacologia, 21:209-262.
DAVIS, G. M., M. MAZURKIEWICZ and M. MANDRACCHIA. 1982. Spurwinkia: Morphology, Systematics, ecology of a new genus of North American Marshland Hydrobiidae (Mollusca: Gastropoda). Proc. Acad. Nat. Sci. of Phila., 134-143-177.
DAVIS, J. R. 1983. An additional record of living Orygoceras (Hydrobiidae) from Texas. Nautilus 97:112-113.
DEACON, J. E. and W. L. MINCKLEY. 1974. Desert fishes. Pp. 385-488. In: Desert Biology, Vol. 11 (G. W. Brown, Jr., Ed.), Academic Press, NY.
FRETTER, V. and A. GRAHAM. 1962. British Prosobranch Molluscs. Ray Soc., London, 755p
FULLINGTON, R. W. 1978. The Recent and Fossil freshwater gastropod fauna of Texas. Unpubl. Ph.D. Dissert., N. Tex. State Univ., Denton.
HERSHLER, R. 1983. The systematics and evolution of the hydrobioid snails (Gastropoda: Rissoacea) of the Cuatro Ciénegas Basin, Coahuila, Mexico. Malacologia. In Press.
HERSHLER, R. and G. M. DAVIS. 1980. The morphology of Hydrobia truncate (Gastropoda: Hydrobiidae): relevance to systematics of Hydrobia. Biol. Bull. 158:195-219.
HOLSINGER, J. R. 1973. Two new species of the subterranean amphipod genus Mexiweckelia (Gammaridae) from Mexico and Texas, with notes on the origin and distribution of the genus. Pp. 1-12. In: Studies on the cavernicole fauna of Mexico and adjacent regions. Assoc. Mexican Cave Stud. Bull. 5.
HOLSINGER, J. R., and G. LONGLEY. 1980. The subterranean amphipod crustacean fauna of an artesian well in Texas. Smithsonian Contrib. Zool. No. 308. 62 pp.
LABOUNTY, J. F. 1974. Materials for the revision of cichlids from Northern Mexico and Southern Texas, USA (Perciformes: Cichlidae). Unpubl. Ph.D. Disserat., Ariz. State Univ., Tempe.
LELOUP, E. 1953. Gasteropodes. Exploration Hydrobiologique du Lac Tanganyika (1946-1947). Institut Royal des Sciences Naturelles de Belgigue, Bruxelles 3:1-273.
MAGNIEZ, G. 1972. Duex Stenasellidae cavernicoles nouveaux de l'Amerique central: Mexistenasellus parzefalli n. sp. et Mexistenasellus wilkensi n. sp. (Crustacea, Isopoda, Asellota). Internat. J. Spelcol. 4:19-31.
MILLER, R. R. 1978 (1974). Composition and derivation of the endemic fish fauna of the Chihuahuan Desert Region. Pp. 365-382. In: Trans. Symp. Biol. Resour. Chih. Des. Reg., U.S. and Mexico. Nat. Park Serv. Trans. Proc. Ser. 3.
MINCKLEY, W. L. 1969. Environments of the Bolsón of Cuatro Ciénegas, Coahuila, Mexico. Univ. Tex. (El Paso) Sci. Ser. 2:1-65.
MINCKLEY, W. L. 1978 (1974). Endemic fishes of the Cuatro Ciénegas Basin, northern Coahuila, Mexico. Pp. 383-404. In: Trans. Symp. Biol. Res. Chih. Des. Reg., U.S. and Mexico. Nat. Park Serv. Trans. Proc. Ser. 3.
MINCKLEY, W. L, and G. A. COLE. 1968. Preliminary limnologic information on waters of the Cuatro Ciénegas Basin, Coahuila, Mexico. Southwest. Nat. 13:421-43 1.
MORRISON, J. P. E. 1949. The cave snails of eastern North America. Bull. Amer. Malacol. Union 1948: 13-15.
PARODIZ, J. J. 1969. The Tertiary non-marine Mollusca of South America. Ann. Carnegie Mus. 40:1-242.
PILSBRY, H. A. and J. BEQUAERT. 1927. The aquatic mollusks of the Belgian Congo, with a geographical and ecological account of Congo malacology. Bull. Amer. Mus. Nat. Hist. 53:69-202.
PILSBRY, H. A., and J. H. FERRISS. 1906. Mollusca of the Southwestern states 11. Pp. 123-175. In: Proc. Acad. Nat. Sci. of Phila.
PONDER, W. F. 1983. Review of the genera of the Barleeidae (Mollusca: Gastropoda: Rissoacea). Rec. Aust. Mus. In Press.
PRATT, W. L. 1977. Hydrobiid snails of the Moapa warm spring complex, Nevada. West. Soc. Malacol., Ann. Rep. 1977 10:7.
RUSSELL, R. H. 1971. Mollusca of Fish Springs, Juab County, Utah: rediscovery of Stagnicola pilsbryi (Hemphill, 1890). Gr. Bas. Nat., 31:223-236.
STANLEY, S. M. 1979. Macroevolution: Pattern and Processes. W. H. Freeman and Co., San Francisco. 132 pp.
TAYLOR, D. W. 1966a. A remarkable snail fauna from Coahuila, Mexico. Veliger, 9.152-228.
TAYLOR, D. W. 1966b. Summary of North American Blancan non-marine molIusks. Malacologia, 4:1-171.
TAYLOR, D. W. 1974. The tertiary gastropod Orygoceras found living. Archiv fur Molluskenkunde, 107:93-96.
TAYLOR, D. W, and W. L. MINCKLEY. 1966. New world for biologists. Pac. Disc., 19:18-22.
THOMPSON, F. G. 1968. The aquatic snails of the family Hydrobiidae of peninsular Florida, Univ. Fla. Press, Gainesville. 268 pp.
THOMPSON, F. G. 1979. The systematic relationships of the hydrobiid snail genus Nymphophilus Taylor 1966 and the status of the subfamily Nymphophilinae. Malacol. Rev., 12.41-50.
THOMPSON, F. G and J. E. McCALEB. 1978. A new freshwater snail from a spring in Eastern Alabama. Amer. Midl. Nat., 100:350-357.
VERMEIJ, G.J. and A. P. COVICH. 1978. Coevolution of freshwater gastropods and their predators. Amer. Nat., 112:833-843.
YONGE, C. M. 1938. The prosobranchs of Lake Tanganyika. Nat., 142:464-466.